Железистые и железисто-фиброзные полипы эндометрия
Функциональная диагностика и УЗИ
Карпочев
Максим Викторович
Баранова
Юлия Викторовна
Фроловская
Людмила Викторовна
Базарнова
Евгения Васильевна
Басаков
Кирилл Сергеевич
Рентгенология
Кивасев
Станислав Александрович
Фельдшеров
Михаил Викторович
 отдел. лучевой диагностики
отдел. лучевой диагностикиНечаев
Валентин Александрович
Басарболиев
Алексей Викторович
Тихонова
Валерия Сергеевна
Мухин
Андрей Андреевич
Грибкова
Мария Сергеевна
Крысанова
Александра Владимировна
Шульц
Евгений Игоревич
Звездина
Дарья Максимовна
Терапия
Горбачева
Елена Владимировна
Карданова
Ольга Дмитриевна
Шашкова
Татьяна Валерьевна
 диагностики
диагностикиКомиссаренко
Ирина Арсеньевна
Слащева
Ольга Михайловна
Шаповалова
Вероника Сергеевна
Кускунова
Евгения Александровна
Пульмонология
Физиотерапия
Родина
Елена Вячеславовна
Хан
Иннокентий Евгеньевич
Кардиология
Горбачева
Елена Владимировна
Карданова
Ольга Дмитриевна
Шашкова
Татьяна Валерьевна
 диагностики
диагностикиКомиссаренко
Ирина Арсеньевна
Аудиология и слухопротезирование
Паукова
Марина Владимировна
Колтышева
Екатерина Борисовна
Левина
Юлия Викторовна
Неврология и мануальная терапия
Замерград
Максим Валерьевич
Небожин
Александр Иванович
Иванова
Татьяна Андреевна
Екушева
Евгения Викторовна
Толстенева
Александра Игоревна
Лабораторные услуги
Дерматология и трихология
Телешева
Ирина Вадимовна
Массаж
Ермуш
Станислав Геннадьевич
Эндокринология
Бахтеева
Ирина Владимировна
Аллергология-иммунология
Стационар
Гнелица
Николай Викторович
Добролюбов
Евгений Евгеньевич
Флебология
Морозов
Константин Моисеевич
Голованчук
Лилия Петровна
Косметология
Телешева
Ирина Вадимовна
Гепатология
Комиссаренко
Ирина Арсеньевна
Гинекология
Душкина
Ирина Александровна
Горский
Сергей Леонидович
Егорова
Елена Анатольевна
Афанасьев
Максим Станиславович
Баранова
Юлия Викторовна
Фроловская
Людмила Викторовна
Проктология
Мормышев
Вячеслав Николаевич
Петрук
Максим Николаевич
Педиатрия
Варенкова
Ольга Владимировна
Поддо
Галина Николаевна
Небожин
Александр Иванович
Маркина
Елена Александровна
Строк
Ирина Викторовна
Телешева
Ирина Вадимовна
Болучевский
Дмитрий Николаевич
Фроловская
Людмила Викторовна
Малышева
Ольга Дмитриевна
Шафоростова
Екатерина Васильевна
Медведева
Анна Олеговна
Толстенева
Александра Игоревна
Маргиева
Диана Анатольевна
Цибиков
Илья Владимирович
Криворотько
Михаил Сергеевич
Эндоскопия
Мардачев
Олег Александрович
Хайдурова
Татьяна Константиновна
Центр травматологии и ортопедии
ЛОР (оториноларингология)
Варенкова
Ольга Владимировна
Марковская
Наталья Геннадьевна
Харина
Дарья Всеволодовна
Мирошниченко
Андрей Петрович
Коршунова
Наталья Александровна
Малышева
Ольга Дмитриевна
Джафарова
Марьям Зауровна
Гастроэнтерология
Комиссаренко
Ирина Арсеньевна
Урология-андрология
Долженок
Андрей Николаевич
Болучевский
Дмитрий Николаевич
Маргиева
Диана Анатольевна
Шамов
Денис Алексеевич
Стоматология. Терапия
Терапия
Кужанова
Аннета Магометовна
Орлова
Елизавета Сергеевна
Бабкина
Екатерина Сергеевна
Хирургия
Горский
Сергей Леонидович
Мирошниченко
Андрей Петрович
Трофимова
Ольга Викторовна
Малышева
Ольга Дмитриевна
Медведева
Анна Олеговна
Туманов
Андрей Борисович
Кипарисов
Владислав Борисович
Голованчук
Лилия Петровна
Джафарова
Марьям Зауровна
Боблак
Юлия Александровна
Психотерапия
Поддо
Галина Николаевна
Офтальмология
Миронкова
Елена Александровна
Паршунина
Ольга Алексеевна
 лазерный хирург
лазерный хирургВерещагин
Лев Владиславович
Центр головокружения и нарушения равновесия
Паукова
Марина Владимировна
Замерград
Максим Валерьевич
Колтышева
Екатерина Борисовна
Мельников
Олег Анатольевич
Иванова
Татьяна Андреевна
Травматология и ортопедия
Дедов
Сергей Юрьевич
Циркин
Сергей Сергеевич
Степанов
Андрей Борисович
Герасимов
Денис Олегович
Цибиков
Илья Владимирович
Криворотько
Михаил Сергеевич
МРТ
Кивасев
Станислав Александрович
Фельдшеров
Михаил Викторович
 отдел. лучевой диагностики
отдел. лучевой диагностикиНечаев
Валентин Александрович
Басарболиев
Алексей Викторович
Тихонова
Валерия Сергеевна
Мухин
Андрей Андреевич
Грибкова
Мария Сергеевна
Крысанова
Александра Владимировна
Шульц
Евгений Игоревич
Звездина
Дарья Максимовна
Компьютерная томография
Кивасев
Станислав Александрович
Фельдшеров
Михаил Викторович
 отдел. лучевой диагностики
отдел. лучевой диагностикиНечаев
Валентин Александрович
Басарболиев
Алексей Викторович
Тихонова
Валерия Сергеевна
Мухин
Андрей Андреевич
Грибкова
Мария Сергеевна
Крысанова
Александра Владимировна
Шульц
Евгений Игоревич
Звездина
Дарья Максимовна
Маммография
Кивасев
Станислав Александрович
Крысанова
Александра Владимировна
Басаков
Кирилл Сергеевич
Денситометрия
Кивасев
Станислав Александрович
Фельдшеров
Михаил Викторович
 отдел. лучевой диагностики
отдел. лучевой диагностикиНечаев
Валентин Александрович
Басарболиев
Алексей Викторович
Тихонова
Валерия Сергеевна
Мухин
Андрей Андреевич
Грибкова
Мария Сергеевна
Звездина
Дарья Максимовна
Нефрология
Маркина
Елена Александровна
Центр нефрологии
Детская стоматология
Орлова
Елизавета Сергеевна
Стоматология. Хирургия
Хирургия
Кулиш
Александр Александрович
Стоматология. Ортопедия
Богословский
Владимир Александрович
Захарченко
Александр Валериевич
Диагностика COVID-19
Маммология
Басаков
Кирилл Сергеевич
Артемьев
Алексей Александрович
Online-консультация врача от 1490 ₽
Паукова
Марина Владимировна
Варенкова
Ольга Владимировна
Карданова
Ольга Дмитриевна
Марковская
Наталья Геннадьевна
Шашкова
Татьяна Валерьевна
 диагностики
диагностикиПоддо
Галина Николаевна
Комиссаренко
Ирина Арсеньевна
Левина
Юлия Викторовна
Иванова
Татьяна Андреевна
Миронкова
Елена Александровна
Маркина
Елена Александровна
Циркин
Сергей Сергеевич
Бахтеева
Ирина Владимировна
Строк
Ирина Викторовна
Паршунина
Ольга Алексеевна
 лазерный хирург
лазерный хирургКоршунова
Наталья Александровна
Петрук
Максим Николаевич
Малышева
Ольга Дмитриевна
Медведева
Анна Олеговна
Туманов
Андрей Борисович
Депозитная система
Служба помощи на дому
Слащева
Ольга Михайловна
Коршунова
Наталья Александровна
Горбачев
Евгений Михайлович
Медицинские справки
Стоматология. Имплантология
Имплантология
Кулиш
Александр Александрович
МРТ открытого типа
Кивасев
Станислав Александрович
Фельдшеров
Михаил Викторович
 отдел. лучевой диагностики
отдел. лучевой диагностикиНечаев
Валентин Александрович
Басарболиев
Алексей Викторович
Тихонова
Валерия Сергеевна
Мухин
Андрей Андреевич
Грибкова
Мария Сергеевна
Крысанова
Александра Владимировна
Шульц
Евгений Игоревич
Звездина
Дарья Максимовна
Центр маммологии
Стоматология.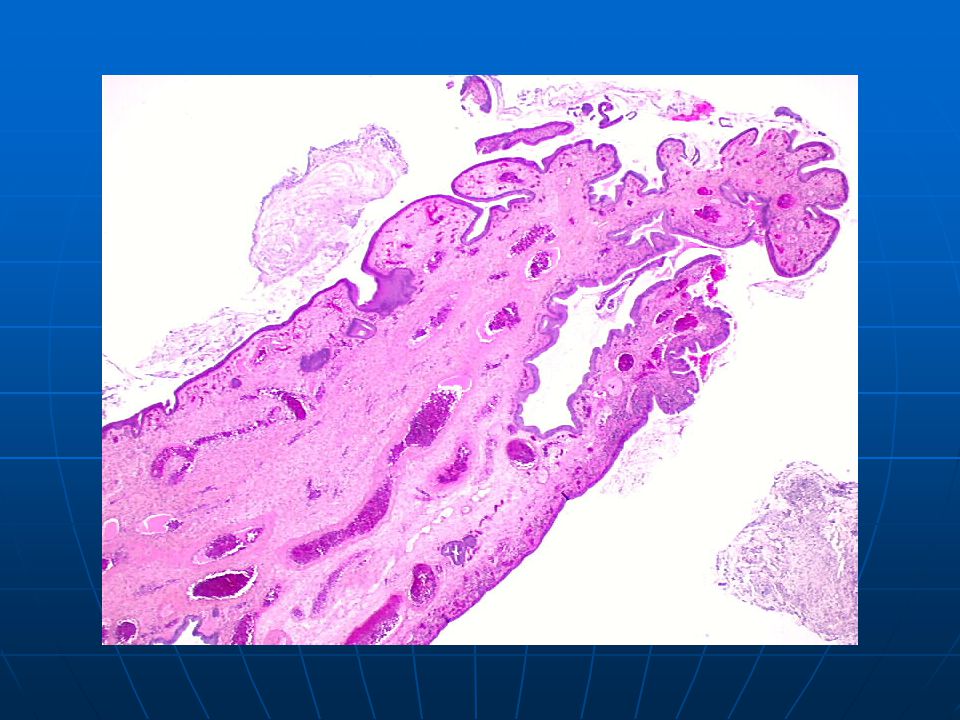 Ортодонтия
Ортодонтия
Шафоростова
Екатерина Васильевна
Миниинвазивные вмешательства при полипах эндометрия и эндоцервикса у пациенток репродуктивного возраста
Бесплодие — одна из актуальных проблем современного общества. Среди супружеских пар детородного возраста частота бесплодия в разных регионах России колеблется от 8 до 19% [1, 2].
В современных условиях отмечается неуклонный рост многих гинекологических заболеваний. Внутриматочная патология (ВМП), в том числе миома матки, гиперпластические процессы в эндометрии, патология цервикального канала, внутриматочные синехии, остается актуальной проблемой гинекологических заболеваний из-за высокой распространенности — у 5—32% женщин репродуктивного возраста [3—7], в том числе у пациенток, использовавших вспомогательные репродуктивные технологии [8].
Внутриматочная патология (ВМП), в том числе миома матки, гиперпластические процессы в эндометрии, патология цервикального канала, внутриматочные синехии, остается актуальной проблемой гинекологических заболеваний из-за высокой распространенности — у 5—32% женщин репродуктивного возраста [3—7], в том числе у пациенток, использовавших вспомогательные репродуктивные технологии [8].
Наиболее часто встречаются полипы эндометрия — у 32%, полипы эндоцервикса — у 22,8%, доля женщин с синехиями составляет 3—13%, с субмукозной миомой — 3%, гиперплазией эндометрия — 0,9%, внутриматочной перегородкой — 0,5%.
Гиперпластические процессы в эндометрии являются предметом изучения многих специалистов: гинекологов, онкологов, эндокринологов, морфологов, что обусловлено в первую очередь риском развития на их фоне рака эндометрия [9—13]. Рост заболеваемости, по данным разных авторов [14, 15], наблюдается в возрастных группах от 40 до 49 лет и от 50 до 59 лет, хотя часто встречается в возрасте до 35 лет и может быть причиной бесплодия.
Основными клиническими проявлениями гиперпластических процессов в эндометрии являются метроррагии, как совпадающие со сроками менструации, так и ациклические. У молодых женщин в 80% случаев гиперпластические процессы в эндометрии ведут к бесплодию, механизмом развития которого является ановуляция или недостаточность лютеиновой фазы, нарушения морфофункционального состояния эндометрия и эндоцервикса [14, 16—18]. Диагностика данных заболеваний позволяет определить тактику ведения больных, выбрать оптимальный метод лечения и у многих пациенток сохранить репродуктивную функцию.
Внедрение в клиническую практику гистероскопии существенно расширило возможности диагностики патологических состояний эндометрия и в то же время позволило при необходимости производить некоторые лечебные манипуляции, а создание минигистероскопов позволило не только проводить исследование в амбулаторных условиях без расширения цервикального канала и использования влагалищных зеркал и пулевых щипцов, но и в ряде случаев одновременно провести лечение.
Цель исследования — повышение эффективности лечения пациенток репродуктивного возраста с полипами эндометрия и эндоцервикса с использованием минигистероскопии.
Материал и методы
В клинике акушерства и гинекологии им. В.Ф. Снегирева Первого Московского государственного медицинского университета им. И.М. Сеченова нами были обследованы и пролечены 90 женщин репродуктивного возраста c полипами эндометрия и эндоцервикса в возрасте от 25 до 46 лет (средний возраст 36,2±0,7 года).
Пациентки были разделены на две группы: 1-я —70 женщин с полипами эндометрия, 2-я — 20 женщин с полипами цервикального канала.
Основными критериями отбора больных для данного исследования являлись полипы эндометрия и эндоцервикса, выявленные на основании результатов функциональных и/или инструментальных методов исследования, репродуктивный возраст пациентки, отсутствие тяжелой соматической патологии.
С целью обследования больных наряду с анализом преморбидного фона, перенесенных и сопутствующих гинекологических заболеваний, особенностей менструальной и генеративной функций, проведены общеклинические, лабораторные исследования, трансвагинальное ультразвуковое сканирование.
Трансвагинальное ультразвуковое исследование осуществляли на ультразвуковом аппарате Toshiba SSH-140A с трансвагинальным конвексным датчиком с частотой 6,5 МГц. Оценивали размеры матки, наличие аномалий развития и деформации матки, структуру миометрия, толщину эндометрия, размеры и структуру яичников, наличие желтого тела и фолликулов.
Проводили лечение с помощью минигистероскопа с каналом для операционных инструментов, диаметром 1,8 мм (5 Fr*), который позволял применять лазерные кварцевые световоды. Для инстилляции жидкости с целью регуляции скорости потока жидкости и давления использовали электрическую помпу.
Процедура проводилась в раннюю фазу пролиферации (6—10-й день менструального цикла), так как именно в эти дни состояние полости матки оптимально для исследования.
Во время минигистероскопии осуществляли осмотр стенок полости матки, оценивали состояние эндометрия, слизистой оболочки цервикального канала и устьев маточных труб, проводили забор материала для гистологического исследования. Полипэктомию в цервикальном канале и полости матки осуществляли при помощи твердотельного лазера на алюмо-иттриевом гранате с неодидом (Nd-YAG) с длиной волны 1,06 мкм в непрерывном режиме на базе установки Medilas 5010 Fibertom.
Полипэктомию в цервикальном канале и полости матки осуществляли при помощи твердотельного лазера на алюмо-иттриевом гранате с неодидом (Nd-YAG) с длиной волны 1,06 мкм в непрерывном режиме на базе установки Medilas 5010 Fibertom.
Результаты и обсуждение
Проведен анализ результатов 90 минигистероскопий. В 1-й группе у 60 (66%) пациенток обнаружены единичные полипы эндометрия, у 10 (11%) — множественные полиповидные образования. Во 2-й группе у 14 (15%) пациенток обнаружены единичные полипы цервикального канала (рис. 1), у 6 (7%) — два полиповидных образования.
Рис. 1. Полип цервикального канала. Минигистероскопия.
Основной жалобой больных была жалоба на отсутствие беременности — у 65 (72%) пациенток. У 36 (40%) женщин бесплодие продолжалось от 1 года до 5 лет и у 29 (32%) больных — от 5 до 10 лет; 15 (17%) женщин предъявляли жалобы на маточные кровотечения, совпадающие со сроками менструации, межменструальные кровянистые выделения из половых путей.
При изучении акушерско-гинекологического анамнеза установлено, что беременности были у 74 (82%) пациенток, из них родами закончились у 45 (50%). Одни роды в анамнезе были у 31 (34%) женщины, двое родов — у 14 (16%).
В 55 (61%) наблюдениях имелось искусственное прерывание беременности, Самопроизвольные аборты с последующим выскабливанием стенок полости матки произошли у 31 (34%) пациентки, среди них у 5 (6%) были поздние самопроизвольные аборты.
В 50 (55%) наблюдениях было произведено экстракорпоральное оплодотворение (ЭКО) и перенос эмбрионов (ПЭ), среди них у 21 (23%) пациентки наступила беременность, лишь у 5 (6%) из них беременность завершилась родами. У 75 (83%) пациенток в анамнезе были внутриматочные хирургические вмешательства. Частота различных внутриматочных хирургических вмешательств у пациенток репродуктивного возраста представлена на рис. 2.
Рис. 2. Частота внутриматочных хирургических вмешательств у пациенток репродуктивного возраста. РДВ — раздельное диагностическое выскабливание.
Всем пациенткам с полипами эндометрия и цервикального канала было произведено оперативное лечение в объеме гистероскопии, лазерной полипэктомии с использованием Nd-YAG-лазерного излучения (рис. 3).
Рис. 3. Лазерная полипэктомия с использованием Nd-YAG-лазерного излучения.
У 76 (84%) пациенток процедура проводилась под внутривенной анестезией без расширения цервикального канала, что позволило минимизировать травматичность и сократить время операции, у 14 (15%) пациенток с единичными полипами цервикального канала — без наложения пулевых щипцов и анестезии.
Все материалы отправлялись на гистологическое исследование. По результатам гистологического исследования в 1-й группе железисто-фиброзные полипы выявлены у 25 (28%) пациенток, железистые полипы — у 42 (47%), фиброзные полипы — у 2 (2%) и у 1 (1%) пациентки — плацентарный полип.
Во 2-й группе железисто-фиброзные полипы установлены у 7 (23%) пациенток, железистые полипы — у 12 (13%), фиброзный полип — у 1 (1%).
При проведении 90 минигистероскопий мы не наблюдали осложнений в виде перфорации стенок матки, кровотечения, тромбоэмболии легочной артерии, отека легких.
Продолжительность операции в среднем составила 10,7±4 мин, средняя кровопотеря — 30,6±0,9 мл. Время пребывания в стационаре в среднем составило 1,4±0,6 дня, средняя продолжительность раннего послеоперационного периода — 2,2±0,5 дня. В послеоперационном периоде проводилась комплексная противовоспалительная, гормональная, ангиопротекторная, антигипоксическая терапия, позволяющая не только избежать рецидивов заболевания, но и восстановить функциональное состояние эндометрия и миометрия.
Для контроля за состоянием полости матки в дальнейшем проводили ультразвуковое исследование органов малого таза и в ряде случаев офисную гистероскопию в амбулаторных условиях. У 65 (72%) пациенток была произведена офисная минигистероскопия спустя 2—3 менструальных цикла после вмешательства, при которой у всех пациенток внутриматочной патологии выявлено не было. У всех пациенток при дальнейшем ультразвуковом контроле в течение года рецидивов полипов не отмечали.
Исходы беременности, наступившей после проведенного лечения, у пациенток обеих групп приведены на рис. 4.
Рис. 4. Исходы беременности, наступившей после лечения, у пациенток обеих групп. а — самопроизвольно наступившая беременность; б — беременность, наступившая после применения программ ЭКО и ПЭ.
Беременность после лечения наступила у 43 пациенток: у 26 пациенток в 1-й группе, у 17 — во 2-й группе. При этом самопроизвольно беременность наступила у 16 пациенток (см. рис. 4, а), а после применения программ ЭКО и ПЭ — у 27 (см. рис. 4, б).
Таким образом, комплексный миниинвазивный метод лечения пациенток репродуктивного возраста с внутриматочной патологией (полипы эндометрия и эндоцервикса), включающий лазерохирургическое вмешательство, гормональную, этиотропную, противовоспалительную и антибактериальную терапию, является высокоэффективным методом, позволяющим не только избежать рецидивов заболевания, но и восстановить функциональное состояние эндометрия и миометрия, что является основным при лечении больных с бесплодием. Преимуществом минигистероскопии является сочетание диагностической и лечебной процедуры, объединение их в единую методику, в которой хирургическая часть «встроена» в диагностическое обследование, позволяет не только «увидеть», но и одновременно устранить выявленную патологию. Необходимо отметить, что в случае использования такого дорогостоящего метода лечения бесплодия, как ЭКО, полость матки должна быть «идеальной». Минигистероскопия, став рутинным методом исследования и лечения, в совокупности с другими диагностическими ресурсами позволит этого добиться.
Минигистероскопия — малоинвазивная, технически простая и высокоинформативная процедура, в некоторых случаях не требующая обезболивания, может применяться в качестве скринингового метода обследования у женщин с бесплодием, а также в ряде случаев может использоваться как оперативное вмешательство без дополнительной травматизации цервикального канала и шейки матки. Включение минигистероскопии в обследование пациенток с бесплодием на этапе подготовки к ЭКО позволяет своевременно диагностировать и в ряде случаев устранять внутриматочную патологию, что дает возможность улучшить состояние эндометрия и повысить эффективность преодоления женского бесплодия.
Особенности экспрессии молекулярно-биологических маркеров в железисто-фиброзных полипах эндометрия у женщин пожилого и старческого возраста
Цель исследования. Изучение экспрессии маркеров пролиферации в клетках железисто-фиброзных полипов эндометрия (ЖФПЭ) для уточнения патогенетических аспектов гиперпластических процессов в эндометрии у пациенток пожилого и старческого возраста.ГБОУ ВПО Московский государственный медико-стоматологический университет им. А.И. Евдокимова Минздрава России; ФГБУ Клиническая больница Управления делами Президента РФ, Москва
Материал и методы. Обследованы 288 пациенток пожилого и старческого возраста c подозрением на патологический процесс в эндометрии по результатам ультразвукового скрининга, которым была выполнена гистероскопия, биопсия эндометрия с последующим морфологической верификацией. Иммуногистохимическое исследование выполнено 9 пациенткам с ЖФПЭ в постменопаузе, группу сравнения составили 10 пациенток в репродуктивном возрасте.
Результаты. У женщин в постменопаузальном периоде экспрессия рецепторов эстрогенов и прогестерона ядрами эпителиальных и стромальных клеток ЖФПЭ, статистически достоверно ниже по сравнению с репродуктивным возрастом. Экспрессия ингибитора апоптоза bcl-2 в эпителиальных и стромальных клетках более выражена, а экспрессия индуктора апоптоза, антигена Вах, наоборот, значительно подавлена. Также выражена экспрессия АЦР450, в основном в эпителии желез,.
Заключение. Обнаружены принципиальные молекулярно-биологические отличия ЖФПЭ у женщин в постменопаузе от таковых в репродуктивном периоде, свидетельствующие о меньшей зависимости полипов от влияния эстрогенов и прогестерона и повышенном риске неопластической трансформации.
гиперпластические процессы в эндометрии
железисто-фиброзные полипы эндометрия
экспрессия рецепторов прогестерона и эстрогенов
апоптоз
- Стрижова Н.В., Сухих Г.Т., Соболева Г.М., Холодова Ж.Л., Рейснер Е.А. Сравнение сывороточной активности матриксной металлопротеиназы-2 у больных с железистой и железисто-кистозной гиперплазиями эндометрия в перименопаузе. Проблемы репродукции. 2008; Спецвыпуск: 312-3.
- Торчинов А.М., Умаханова М.М., Фидарова Т.В., Габараева М.Р. Пролиферативная активность железистого эпителия полипов эндометрия в период пери- и постменопаузы. В кн.: Материалы IX Всероссийского научного форума «Мать и дитя». М.; 2007: 540-1.
- Адамян Л.В., Спицын В.А., Андреева Е.Н. Генетические аспекты гинекологических заболеваний. М.: ГЭОТАР-Медиа; 2008. 192с.
- Бохман Я.В. Руководство по онкогинекологии. СПб.: Фолиант; 2002: 195-229.
- Бреусенко В.Г., Савельева Г.М., Голова Ю.А., Мишиева О.И. Лечение гиперпластических процессов эндометрия в периоде постменопаузы. Акушерство и гинекология. 2009; 4: 19-23.
- Cавельева Г.М., Бреусенко В.Г., Голова Ю.А., Мишиева О.И., Ивановская Т.Н. Гиперпластические процессы эндометрия в пре- и постменопаузе. Методы терапии. Онкогинекология. 2012; 1: 43-6.
- Сметник В.П., ред. Медицина климактерия. М.: Литера; 2006: 187-216.
- Табакман Ю.Ю. Рак эндометрия. Руководство для врачей. М.: Практическая медицина; 2009: 134-7.
- Anderson A.S., Caswell S. Obesity management –an opportunity for cancer prevention. Surgeon. 2009; 7(5): 282-5.
- Burbos N., Musonda P., Giarenis I., Shiner A.M., Giamougiannis P., Morris E.P., Nieto J.J.. Predicting the risk of endometrial cancer in postmenopausal women presenting with vaginal bleeding: the Norwich DEFAB risk assessment tool. Br. J. Cancer. 2010; 102(8): 1201-6.
- Рымашевский А.Н., Воробьев С.В., Андрющенко Ю.А. Клиническая эффективность комбинированной оперативной и гормонально-метаболической терапии полипов эндометрия у женщин с ожирением в постменопаузе. Современные проблемы науки и образования. 2011; 3.
- Wolfman W., Leyland N., Heywood M., Singh S.S., Rittenberg D.A., Soucy R. et al.; Society of Obstetricians and Gynaecologists of Canada. Asymptomatic endometrial thickening. J. Obstet. Gynaecol. Can. 2010; 32(10): 990-9.
- Ашрафян Л.А., Киселев В.И. Опухоли репродуктивных органов (этиология и патогенез). М.: Димитрейд График Групп; 2008. 216с.
- Девятовская А.Г. Применение трехмерной эхографии в мониторинге лечения гиперпластических процессов эндометрия: дисс. … канд. мед. наук. М.; 2009. 117с.
- Саркисов С.Э., Хитрых О.В., Куковенко Е.М. Отдаленные результаты и оптимизация тактики лечения полипов эндометрия в постменопаузе. Акушерство и гинекология. 2009; 5: 44-8.
- Agostini A., Collette E., Provansal M., Estrade J.P., Blanc B., Gamerre M. Guidelines and accuracy of office hysteroscopy and endometrial biopsy. J. Gynecol. Obstet. Biol. Reprod. 2008; 37( Suppl.8): S343-8.
- Schmidt T., Breidenbach M., Nawroth F., Mallmann P., Beyer I.M., Fleisch M.C., Rein D.T. Hysteroscopy for asymptomatic postmenopausal women with sonographically thickened endometrium. Maturitas. 2009; 62(2): 176-8.
- AAGL practice report: practice guidelines for the diagnosis and management of endometrial polyps. J. Minim. Invasive Gynecol. 2012; 19(1): 3-10.
- Brand A.H. The woman with postmenopausal bleeding. Aust. Fam. Physician. 2007; 36(3): 116-20.
- Renaud M.C., Le T., Bentley J., Farrell S., Fortier M.P., Giede C. et al. Epidemiology and investigations for suspected endometrial cancer. J. Obstet. Gynaecol. Can. 2013; 35(4): 380-3.
Поступила 19.06.2015
Принята в печать 26.06.2015
Зайратьянц О.В., д.м.н., профессор, зав. кафедрой патологической анатомии МГМСУ им. А.И. Евдокимова Минздрава России. Адрес: 127473, Россия, Москва, ул. Делегатская, д. 20, стр. 1
Сонова М.М., д.м.н., доцент кафедры репродуктивной медицины и хирургии МГМСУ им. А.И. Евдокимова Минздрава России. Адрес: 127473, Россия, Москва, ул. Делегатская, д. 20, стр. 1
Мельникова Н.С., акушер-гинеколог гинекологического отделения ФГБУ Клиническая больница Управления делами Президента РФ. Адрес: 107150, Россия, Москва, ул. Лосиноостровская, д. 45. Телефон: 8 (916) 151-14-59. E-mail: [email protected]
Балицкий Е.В., заведующий патологоанатомическим отделением ФГБУ Клиническая больница Управления делами Президента РФ. Адрес: 107150, Россия, Москва, ул. Лосиноостровская, д. 45
Для цитирования: Адамян Л.В., Зайратьянц О.В., Сонова М.М., Мельникова Н.С., Балицкий Е.В. Особенности экспрессии молекулярно-биологических маркеров в железисто-фиброзных полипах эндометрия у женщин пожилого и старческого возраста. Акушерство и гинекология. 2015; 11:
Полип эндометрия – виды, причины возникновения и методы лечения
Полип эндометрия представляет собой доброкачественное образование в полости матки. Если в органе есть много таких образований, то говорят о полипозе матки. Новообразования растут из базального внутреннего слоя эндометрия. Им страдают женщины как репродуктивного возраста, так и пребывающие в менопаузе. При полипозе матки происходит гиперплазия её слизистой оболочки. Под влиянием мутагенных факторов полипы могут малигнизироваться, то есть, перерождаться в злокачественное новообразование.
Разновидности полипов
Прежде всего, поговорим о том, из каких клеток состоят полипы и каково их строение. Полип обычно состоит из клеток эпителия. Он имеет такие части, как тело и ножка. Различают следующие виды полипов:
• железистые;• фиброзные;
• железисто-фиброзные;
• аденоматозные.
Железистый полип эндометрия, лечение которого проводят в гинекологии, состоит из основы и железистого эпителия. Фиброзный полип эндометрия составляют клетки соединительной ткани. В состав железисто-фиброзных полипов входят как соединительнотканные элементы, так и клетки железистого эпителия. Основными элементами аденоматозных полипов составляет также железистый эпителий, но при гистологическом исследовании в них находят клетки, имеющие признаки атипии.
Этиологические факторы образования полипов
Полипоз матки – это мультифакторное заболевание, которое развивается под воздействием множества причин. Образование полипов слизистой оболочки матки происходит вследствие таких моментов:
• диспропорция между гормонами прогестерон и эстрогены;• хроническая травматизация эндометрия внутриматочной спиралью;
• частые диагностические выскабливания либо прерывания беременности;
• роды, при которых не полностью удаляется плацента, а также выкидыши и частые аборты;
• некоторые соматические заболевания, сопровождающиеся нарушением гормонального фона;
• снижение специфических и не специфических факторов защиты;
• неуправляемая артериальная гипертензия,
• некоторые инфекционные заболевания и воспалительные процессы органов малого таза.
Симптомы полипов эндометрия
Далеко не всегда женщина может заподозрить, что у неё имеется гиперплазия или полип эндометрия. Полипы небольшого размера никак себя не проявляют. Женщина может обратиться к гинекологу по поводу каких-то жалоб или прийти на профилактический осмотр, и он обнаружит полип. Иногда эту паталогию выявляют на УЗИ. Однако в большинстве случаев при полипах матки можно обнаружить достаточное количество патогномоничных симптомов, которые позволяют заподозрить неладное.
Каковы же наиболее характерные симптомы полипов эндометрия? Прежде всего, женщина жалуется на наличие схваткообразных болей в нижних отделах живота, которые усиливаются во время коитуса или появляются сразу же после полового акта. У пациенток репродуктивного возраста с неизменённым циклом увеличивается интенсивность и объём выделений во время менструации.
Меноррагии довольно часто приводят к анемии. Женщина чувствует себя постоянно усталой, её беспокоят головные боли, общая слабость. Кожа и видимые слизистые оболочки становятся бледными, под глазами можно увидеть тёмные круги. Также начинают ломаться ногти и выпадать волосы. Гипотония и тахикардия приводит к головокружениям и коллаптоидным состояниям.
У женщин, пребывающих в постменопаузе, могут один или два раза появится выделения из половых путей, на которые она не обращает никакого внимания. Если полип приобретает большой размер, то могут появиться бели. После коитуса возможны скудные выделения с примесью крови.
Полип эндометрия и беременность
Беременность при полипе эндометрия матки всегда проблемная. Женщины, страдающие полипозом матки, не могут зачать. Если же и произойдёт зачатие, то вряд ли эмбрион сможет имплантироваться в патологически изменённую слизистую матки. Ну а в том случае, когда беременность всё же наступает, то высока угроза выкидыша.
Достаточно часто женщины расценивают кровотечение, которые возникает вследствие полипа, как угрозу выкидыша на раннем сроке. В любом случае женщине при наличии маточных кровотечений следует обратиться к специалисту. Ясно одно, что в любом случае, при наличии маточных кровотечений женщине следует обратиться к гинекологу.
После удаления полипа эндометрия беременность возможна. Лечение полипа эндометрия народными средствами следует проводить только при наличии новообразований небольшого размера. После такого лечения шансы забеременеть увеличиваются.
Методы диагностики полипа эндометрия
Прежде всего, гинеколог, к которому женщина обратилась на консультацию, должен опросить её. После сбора анамнеза проводят обследование пациентки, включающее бимануальный гинекологический осмотр. Далее женщину смотрят в зеркалах. Во время такого обследования можно обнаружить полипы эндометрия, которые расположены в шейке цервикального канала. В дальнейшем применяют следующие методы исследования:
• гистероскопию;• эхоскопию матки (УЗИ) с применением вагинального датчика;
• биопсию с последующим гистологическим исследованием препарата;
• КТ и МРТ (по показаниям).
Гистероскопия – это диагностический метод, который выполняется под общим обезболиванием. Врач вводит в полость матки оптическую камеру, и с помощью световолоконной оптики осматривает эндометрий. Во время этой манипуляции возможно удаление полипа.
В ряде случаев с помощью гистероскопа берут кусочек ткани из подозрительного участка слизистой матки, который отправляют на гистологическое исследование. Это позволяет исключить злокачественный характер новообразования. Наиболее современным способом взятия материала для исследования эндометрия является пайпель-биопсия. Этот метод самый безболезненный, безопасный и короткий по времени проведения.
Пайпель представляет собой специальную пластиковую трубку, тонкую в диаметре, которая имеет небольшой поршень. С его помощью этого осуществляется всасывание эндометрия для предстоящего исследования материала. Вся процедура длится не более тридцати секунд.
Полип матки можно увидеть во время ультразвукового исследования, которое выполняют, используя вагинальный датчик. Это позволяет поставить предварительный диагноз. Компьютерную и магнитно-резонансную томограмму выполняют при множественных полипах.
Методы лечения полипоза эндометрия
В большинстве случаев для лечения полипов эндометрия выполняется операция. Полип матки можно удалить во время гистероскопии. Врач производит удаление паталогии и производит выскабливание слизистой матки под контролем камеры гистероскопа. Если же он обнаружит полип большого размера, то может его просто открутить. После того, как удалили полип эндометрия, полученный материал отправляют в гистологическую лабораторию. Производят прижигание места крепления полипа либо жидким азотом, либо электрокоагулятором.
Можно также выполнить лазерную коагуляцию или криодеструкцию полипа. После лечения полипа эндометрия, на третий день после его удаление, выполняют контрольную эхоскопию матки. А какую тактику выбрать в дальнейшем, врач определит, получив результаты гистологического исследования.
В случае, когда на гистологии обнаружится фиброзный полип эндометрия, лечение заканчивают. Если выяснится, что у пациентки фиброзно-железистый полип эндометрия, лечение назначают с целью нормализации гормонального фона. При наличии железистых и фиброзно-железистых новообразований пациентке любого возраста следует назначить гормональные препараты.
Женщинам, у которых возраст не превышает тридцати лет, назначают для лечения полипов эндометрия комбинированные оральные контрацептивы. Пациенткам после тридцати лет следует назначить гестагены. Если женщина в течение ближайших пяти лет не планирует беременность, ей целесообразно поставить гормональную спираль «Мирена».
Более жёстко подходят к лечению аденоматозного полипа. Если пациентка пребывает перед менопаузой либо в менопаузе, то ей предлагают выполнить экстирпацию (удаление) матки. При подозрении на озлокачествление полипа или дисгормональных расстройствах, следует удалить матку с яичниками. Если же женщина находится в репродуктивном возрасте и планирует беременность, то оперативное вмешательство следует отложить и попробовать провести гормональную терапию.
В послеоперационном периоде для профилактики инфекционных осложнений с профилактической целью назначают антибиотики. В течение десяти дней женщина должна придерживаться диеты: употреблять в пищу низкокалорийные продукты.
Народное лечение полипа эндометрия
Часто у женщин возникает вопрос: а можно ли лечить полип эндометрия народными средствами. В принципе, это возможно. Но вначале нужно обратиться к гинекологу, пройти соответствующее обследование, выполнить гистологическое исследование. Только после этого вместе с врачом следует определиться с тактикой лечения.
Каково же народное лечение полипа эндометрия? Народная медицина предлагает множество методов лечения этого заболевания. Так, проводится спринцевание отваром чистотела, зверобоя, календулы, бодяги. Говорят, что от полипов матки помогает отвар болиголова, настои грибов шиитака или чака, которыми женщине тоже следует спринцеваться.
Также целители рекомендуют вводить во влагалище тампоны с чесноком, принимать внутрь отвары разных трав. Полип эндометрия народными средствами можно лечить, но не факт, что лечение будет успешным. Да и не стоит забывать, что полипы эндометрия способны малигнизироваться. Поэтому, лечение следует проводить под наблюдением гинеколога.
Если вы нуждаетесь в диагностике или лечении полипов эндометрия, обращайтесь в «Центр ЭКО» Нальчик. Наши специалисты проведут необходимое обследование и предложат наименее травматичный метод лечения. Мы применяем индивидуальный подход к лечению полипов матки.
Удаление полипа в матке — лечение полипа шейки матки в Москве по доступной цене
Полипом матки называют доброкачественное образование, которое в большинстве случаев формируется на внутренней стенке указанного органа и выступает в его полость. Оно крепится к стенкам матки благодаря наличию тонкой ножки или широкого основания. Размеры полипа могут достигать нескольких сантиметров. Как правило, такие новообразования располагаются в полости матки, но иногда они могут выступать за ее пределы, прорастая через цервикальный канал во влагалище. Так как полипы не состоят из раковых клеток, они обычно не представляют опасность для здоровья и жизни женщины. В большинстве случаев такие образования рассасываются самостоятельно без дополнительного лечения. Если же женщина жалуется на кровотечения или другие осложнения, вызванные появлением полипа, производится его удаление.
Медицинский центр «Медлайн-Сервис» предлагает услуги квалифицированных специалистов, которые помогут Вам заботиться о своем здоровье. Обращаем Ваше внимание на то, что врачи рекомендуют женщинам проходить профилактические осмотры у гинеколога дважды в год. Это позволит вовремя обнаружить патологии и предпринять необходимые меры для лечения. При наличии каких-либо жалоб также не стоит откладывать консультацию со специалистом. В нашем центре проводится раздельное диагностическое выскабливание, направленное на удаление полипов в матке или цервикальном канале. Для диагностики и лечения мы применяем современное оборудование и методы терапии. На стоимость удаления полипов шейки матки влияет их количество и другие факторы, поэтому ее необходимо уточнять при очной консультации с врачом.
Симптомы полипа маткиИногда процесс образования и роста полипа в матке или на ее шейке может протекать без выраженных симптомов. Но зачастую на фоне подобных отклонений наблюдаются нарушение цикла, изменение характера менструаций, патологические кровотечения в период между месячными и после менопаузы. Кроме того, женщины могут жаловаться на дискомфорт и болезненные ощущения во время полового акта. Все эти симптомы учитываются врачом при постановке диагноза, поэтому во время консультации следует обязательно рассказать о них.
Причины появления полипаПолипы могут образоваться у девушек и женщин любого возраста. К причинам их появления зачастую относят гормональные нарушения, воспалительные процессы, а также наличие эрозии шейки матки. Однако точно сказать, почему произошло развитие этой патологии, врачи в большинстве случаев не в состоянии. Некоторые специалисты утверждают, что причина кроется в повышении в организме уровня эстрогена. Также существует предположение о том, что развитие и рост полипов шейки матки увеличивается, если женщина страдает ожирением, гипертонией или принимает некоторые лекарственные препараты.
Диагностика полипа маткиОбычно для постановки точного диагноза при наличии полипов в матке или цервикальном канале стандартного гинекологического осмотра бывает недостаточно. В таких случаях используют более точные диагностические процедуры, например УЗИ органов малого таза, гистероскопию (для определения размеров и количества новообразований, а также взятия биопсии для исследования), метрографию (рентгенологическое обследование с применением контрастного реагента). Кроме того, возможно выполнение диагностического выскабливания. Эти две операции не только являются диагностическими, но и позволяют одновременно произвести лечение при полипе шейки матки. После их выполнения полученный материал отправляется на гистологию для исследования на раковые клетки.
Виды полиповЖелезистые. Эти новообразования состоят из ткани эндометрия и расположенных в ней желез. Обычно они достигают размера не более 1,5 см.
Железисто-фиброзные. Такие полипы состоят из эндометрия и соединительной ткани. Они могут иметь размер 2–2,5 см.
Фиброзные. Эти новообразования состоят из фиброзной ткани повышенной плотности. Чаще всего они встречаются у женщин пожилого возраста.
Аденоматозные. Такие полипы имеют однородную структуру. Они достигают размера до 4 см.
Зачем удалять полип матки?Несмотря на то, что доброкачественные новообразования шейки матки иногда могут не причинять никаких беспокойств, рекомендуется их удаление. Отсутствие лечения при полипах матки и цервикального канала может привести к серьезным последствиям для здоровья женщины. Они могут проявиться в виде гормональных сбоев и нарушения работы желез внутренней секреции. Кроме того, известны случаи, когда доброкачественные образования перерождались в предраковые состояния или вовсе приводили к развитию рака шейки матки. Именно поэтому даже при отсутствии симптомов патологии не откладывайте процедуру удаления полипа матки. В большинстве случаев это простая операция, которая не требует длительной реабилитации. Цена на удаление полипов матки невысокая по сравнению со стоимостью устранения последствий, к которым может привести отсутствие должного лечения.
Способы лечения полиповЛечение полипов шейки матки предполагает их хирургическое удаление. Выбор метода проведения операции зависит от степени распространенности и характера новообразований, а также индивидуальных особенностей организма. Основным методом удаления полипа шейки матки является гистероскопия с последующим выскабливанием. При наличии ярко выраженной ножки новообразования срезают, а затем прижигают место произрастания с применением электрокоагуляции или жидкого азота. Эту процедуру выполняют во избежание рецидивов. После этого делается повторное УЗИ, которое позволяет убедиться в полном удалении полипов. Если в ходе операции не удалось избавиться от всех новообразований, назначается повторное выскабливание.
Удаление полипов шейки матки не требуется, если новообразования имели фиброзный характер. Опасность для здоровья женщины могут представлять железисто-фиброзные и железистые полипы. Их лечение предполагает не только удаление, но и последующую гормональную терапию. Как правило, врачи назначают средства оральной контрацепции или рекомендуют введение внутриматочных гормональных препаратов. Из-за угрозы перерастания в злокачественные клетки удаленный материал отправляется на гистологический анализ. При лечении полипов матки, носящих аденоматозный характер, прибегают к радикальным мерам. Пациенткам, которые не собираются иметь детей, выполняют операцию по удалению матки. Женщинам детородного возраста проводят хирургическое вмешательство и назначают обязательную гормональную коррекцию.
Рецидивы и последующий прогнозПосле лечения при полипах матки пациентки должны некоторое время находиться под наблюдением и регулярно посещать врача. Реабилитация заключается в нормализации цикла и процедурах, выполняемых для избегания рецидивов. К сожалению, операция по удалению полипов не является гарантией того, что они не появятся снова. При этом рецидивы могут сопровождаться приобретением новообразований злокачественной формы. В группу риска в основном входят женщины, которые столкнулись с аденоматозными полипами. Именно поэтому им уделяется особое внимание в процессе реабилитации. В целом последствия при образовании полипов могут быть разными – от гормонального сбоя до бесплодия, поэтому при наличии каких-либо жалоб не откладывайте посещение гинеколога.
Профилактика появления полиповЧтобы не допустить повторного образования полипов, необходимо знать причину возникновения такой патологии и направить все силы на борьбу с ней. Женщинам рекомендуется внимательно следить за менструальным циклом, его периодичностью и характером выделений. При необходимости следует регулярно сдавать анализы на гормоны и принимать назначенные врачом препараты для их регулирования. Помните, что рецидив заболевания может произойти в любой момент, поэтому не забывайте проходить профилактические осмотры и выполняйте все рекомендации специалистов.
Вы можете задать интересующие Вас вопросы по указанному на сайте контактному номеру. Стоимость удаления полипа матки уточняйте у наших специалистов. Записаться к ним Вы можете по указанному телефону в Москве или с помощью специальной формы на сайте.
Полипы эндометрия (внутренней оболочки матки) — это доброкачественные новообразования эндометрия, которые развиваются вследствие разрастания клеток внутреннего эпителиального слоя матки. Они прикрепляются к телу матки с помощью ножки, в которой располагаются сосуды; по ним происходит питание и кровоснабжение этого патологического выроста. Маточные полипы могут быть одиночными, а могут получить множественное распространение. Виды полипов: Полипы в полости матки бывают железистые (состоят из железистого эпителия), железисто-фиброзные (состоят из железистого эпителия и соединительной ткани) и фиброзные (состоят из соединительной ткани). Клетки полипа могут претерпевать изменения: иногда внутри полипа может замедляться кровообращение, могут быть локальные кровоизлиянии, что в общей сложности приводит к отмиранию клеток тела полипа и возникновению воспаления. В этом случае речь идет об аденоматозном полипе, который относят к предраковому состоянию. Отдельную группу составляют плацентарные полипы, которые формируются из остаточных фрагментов плаценты (после родов). Такие образования сопровождаются длительными обильными кровотечениями, что может стать причиной острого воспаления тела матки. Какой именно полип, то есть его гистологический тип, возможно выяснить только после его удаления и проведения гистологического исследования удаленной ткани. Симптомы полипа эндометрия: Полипы небольших размеров могут не вызывать никаких жалоб. В других случаях могут наблюдаться кровянистые выделения из влагалища после окончания месячных (через 2-3 дня) или в середине цикла. Иногда женщина отмечает появление неприятных ощущений во время половых контактов или кровянистые выделения после них. Боли схваткообразного характера и бесплодие характерны при наличии больших полипов (более 2 см в диаметре). Большой полип препятствует имплантации эмбриона. Клиническая картина зависит не только от типа полипа эндометрия (железистый, железисто-фиброзный или фиброзный) и его размеров, но и от общего состояния здоровья женщины, её возраста, наличия других заболеваний, в том числе и хронических. Врач акушер-гинеколог кабинета бесплодия Хасавюртовского отделения РЦОЗСиР Юнусова… — Республиканский центр охраны здоровья семьи и репродукции
Влияние COVID-19 на репродуктивное здоровье мужчин.
Во время пика пандемии мы переживали за наших мам и пап, дедушек и бабушек, так как больше всех подвержены риску заразиться были пожилые люди и люди с хроническими заболеваниями. Но сейчас уже можно говорить, что коронавирус не прошел незаметно для людей репродуктивного возраста.
Исследования подтверждают, что SARS-CoV-2 это гендерно зависимое заболевание и в большинстве случаев тяжелее его переносят мужчины. «Новая коронавирусная инфекция поражает репродуктивные органы и может приводить к бесплодию» — об этом заявил главный внештатный специалист Минздрава по репродуктивному здоровью, директор НИИ урологии им. Н.А.Лопаткина, академик РАН Олег Аполихин.
После первой волны пандемии, мы стали замечать повышение мужского фактора бесплодия у пациентов вступающих в протоколы ЭКО, наши наблюдения показали, что стали снижаться показатели оплодотворения.
Андрологи бьют тревогу, так как после пика пандемии у пациентов значительно ухудшается морфология сперматозоидов, повышается уровень антиспермальных антител, а также увеличивается уровень ДНК — фрагментированных сперматозоидов, а это в свою очередь снижает процент оплодотворения яйцеклеток и процент имплантации эмбрионов.
Мы провели анализ мировых исследований. Самые первые сообщения о влиянии SARS-CoV-2 на мужскую репродуктивную систему стали появляться в апреле 2020 года. В Китае проводились исследования на небольшой группе пациентов, на наличие вируса в эякуляте у пациентов с умеренно выраженным течением болезни и первые данные показывали, что в эякуляте вирус не был обнаружен. Но китайские медики продолжили исследования в группе пациентов с тяжелым течением заболевания и результаты оказались совсем иные. SARS-CoV-2 обнаружили у 16% процентов мужчин исследуемой группы. Т.е. теоретически, сохраняясь в органах репродуктивной системы, вирус может передаваться половым путем, в дополнение к воздушно-капельному.
Читайте продолжение статьи в карусели➡️
#короновирус #ковидибеременность
Гиперпластические процессы эндометрия у женщин с бесплодием: сопоставление результатов ультрасонографических и патоморфологических исследований эндометрия
Кузык Ю. И., Чорненька А. Н.
Львовский национальный медицинский университет имени Данила Галицкого
Тернопольский государственный медицинский университет им. И.Я. Горбачевского
Цель исследования: определение уровня верификации гиперпластических процессов эндометрия (ГПЭ) у женщин с маточным бесплодием на основании сравнения данных ультразвукового и патоморфологического его исследований.
Материалы и методы. 64 пациенткам репродуктивного возраста проводили ультрасонографическое исследование и биопсию или лечебно-диагностическое выскабливание эндометрия.
Результаты. Выделены три группы ГПЭ: полипы – 33 случая; гиперплазия – 15, сочетание железистой гиперплазии с полипом эндометрия – 16. Полипы эндометрия включали: железистые – 7 случаев, железисто-фиброзные с преобладанием железистого компонента – 13 и железисто-фиброзные с преимуществом стромального компонента – 13. Точность верификации железистых полипов – 82% случаев. Железисто-фиброзные полипы эндометрия с преобладанием железистого компонента диагностированы в 82%. Железисто-фиброзные полипы эндометрия с преимуществом стромального компонента были сложными для ультразвуковой верификации. Точность их диагностики составила 50%. Такие характеристики, как узловая форма, интрамуральное расположение, повышенная эхогенность и отсутствие включений были основанием для ошибочной диагностики фибромиомы матки. Гиперплазию эндометрия удалось верифицировать в 97% случаев. Остальные 3% составляли гистологически диагностированные полипы эндометрия, которые не были распознаны при ультразвуковом исследовании (УЗИ), а расценивались как гиперплазия эндометрия. Точность ультразвуковой диагностики группы сочетания гиперплазии эндометрия и полипов достигала 77%. Полипы на фоне гиперплазии эндометрия имели форму фибромиоматозных узлов. Определяющую роль в правильной диагностике сыграла гистероскопия.
Заключение. Сопоставление данных УЗИ и морфологического исследования свидетельствует о высокой точности обнаружения ГПЭ и их морфологической верификации. Однако в ряде случаев ГПЭ УЗИ не позволяет точно определить характер патологических изменений. Поэтому вопросы изучения патоморфологических особенностей ремоделирования эндометрия при ГПЭ остается открытым и требует новых перспективных подходов. Одним из них, исходя из выявленных патоморфологических изменений при ГПЭ, является применение методов исследования кровотока, в частности трансвагинальной цветовой допплерографии, что может позволить повысить точность диагностики ГПЭ.
Ключевые слова: гиперпластические процессы эндометрия, полип эндометрия, гиперплазия эндометрия, ультразвуковая диагностика, патоморфология, репродуктивный возраст.
1. Вдовиченко Ю.П. Патология эндометрия как причина женского бесплодия / Ю.П. Вдовиченко, А.В. Колесник // Здоровье женщины. – 2011. – № 7. – С. 206–207.
2. Герман Д.Г. Полипы эндометрия в репродуктивном возрасте: штрихи к клиническому портрету / Д.Г. Герман // Репродуктивная эндокринология. – 2016. – № 3. – С. 39–43.
3. Гнатко О.П. Поліпи ендометрія і непліддя в жінок пізнього репродуктивного віку / О.П. Гнатко, Н.Г. Скурятіна // Актуальні питання педіатрії, акушерства та гінекології. – 2015. – № 2. – С. 84–87.
4. Давыдов А.И. Атипическая гиперплазия эндометрия: вопросы морфогенеза, классификация, діагностика и лечение / А.И. Давыдов // Вопросы гинекологии, акушерства и перинатологии. – 2009. – № 3. – С. 93–96.
5. Демидов В.Н. Ультразвуковая диагностика в гинекологии / В.Н. Демидов, Б.И. Зыкин. – М.: Медицина, 1990. – 224 с.
6. Кондриков Н.И. Структурно-функциональные основы гиперпластических изменений эндометрия женщин: Автореф. дисс. … д-ра мед. наук: спец. 14.01.01 «Акушерство и гинекология» / Н.И. Кондриков. – М., 1991. – 51 с.
7. Кондриков Н.И. Патология матки / Н.И. Кондриков. – М.: Практическая медицина, 2008. – 176 с.
8. Красильников Р.Г., Абдуллаев Р.Я. Трансвагинальное ультразвуковое исследование и магниторезонансная томография в диагностике гиперпластических процессов эндометрия / Р.Г. Красильников, Р.Я. Абдуллаев // Международный медицинский журнал. – 2013. – № 1. – С. 113–117.
9. Павловская М.А. Современные представления об этиопатогенезе гиперпластических процессов эндометрия в репродуктивном возрасте / М.А. Павловская, Л. В. Гугикова // Медицинские аспекты здоровья женщины. – 2013. – № 3 (67). – С. 28–34.
10. Стрижаков А.Н Доброкачественные заболевания матки / А.Н. Стрижаков, А.И. Давыдов, В.М. Пашков – М.: Медицина, 2011. – 243 с.
11. Цветовая допплерометрия в диагностике гиперпластических процессов и рака эндометрия / Сидорова И.С., Гуриев Т.Д., Капустина И.Н. [и др.] // Медицинская визуализация. – 2001. – № 2. – С. 88–93.
12. Эхографические и морфологические корреляции гиперпластических процессов эндометрия / Озерская И.А., Белоусов М.А., Щеглова Е.А., Агеева М.И. // Ультразвуковая и функциональная диагностика. – 2002. – № 1. – С. 45–50.
13. Comparison of transvaginal ultrasonography and hysteroscopy in the diagnosis of uterine pathologies / Babacan A., Gun I., Kizilaslan C. [et al.] // Int J Clin Exp Med. – 2014. – Vol. 7, № 3. – P. 764–769. PMid:24753775 PMCid:PMC3992420
14. Accuracy of Transvaginal Ultrasonography, Hysteroscopy and Uterine Curettage in Evaluating Endometrial Pathologies / Wanderley M.D., Álvares M.M., Vogt M.F., Sazaki L.M. // Rev Bras Ginecol Obstet. – 2016. – Vol. 38, № 10. – Р. 506–511. https://doi.org/10.1055/s-0036-1593774; PMid:27769088
15. A diagnostically useful histopathologic feature of endometrial polyp: the long axis of endometrial glands arranged parallel to surface epithelium / Kim K.R., Peng R., Ro J.Y., Robboy S.J. // Am J Surg Pathol. – 2004. – Vol. 28. – P. 1057–1062. https://doi.org/10.1097/01.pas.0000128659.73944.f3; PMid:15252313
16. Kumari M., Gupta A.S. A prospective comparative study to evaluate the efficacy of ultrasonography and hysteroscopy and their correlation with the histopathology of endometrium in a case of abnormal uterine bleeding / Kumari M., Gupta A.S. // Int J Reprod Contracept Obstet Gynecol. – 2015. – Vol. 4, № 6. – P. 1926–1935. https://doi.org/10.18203/2320-1770.ijrcog20151288
17. Mamedalieva N., Isenova S. Comprehensive study of the endometrium of women with reproductive disorders and chronic endometritis / N. Mamedalieva, S. Isenova // Medical and Health Science Journal. – 2012. – Vol. 12. – Р. 31–33. https://doi.org/10.15208/mhsj.2012.41
18. New concepts for an old problem: the diagnosis of endometrial hyperplasia / Sanderson P.A., Critchley H.O.D., Williams A.R.W. [et al.] // Reproduction Update. – 2017. – Vol. 23, № 2. – P. 232–254.
19. Treatment of endometrial polyps: a systematic review / Lieng M., Istre O., Qvigstad E. // Acta Obstet Gynecol Scand. – 2010. – Vol. 89. – P. 992–1002. https://doi.org/10.3109/00016349.2010.493196; PMid:20528202
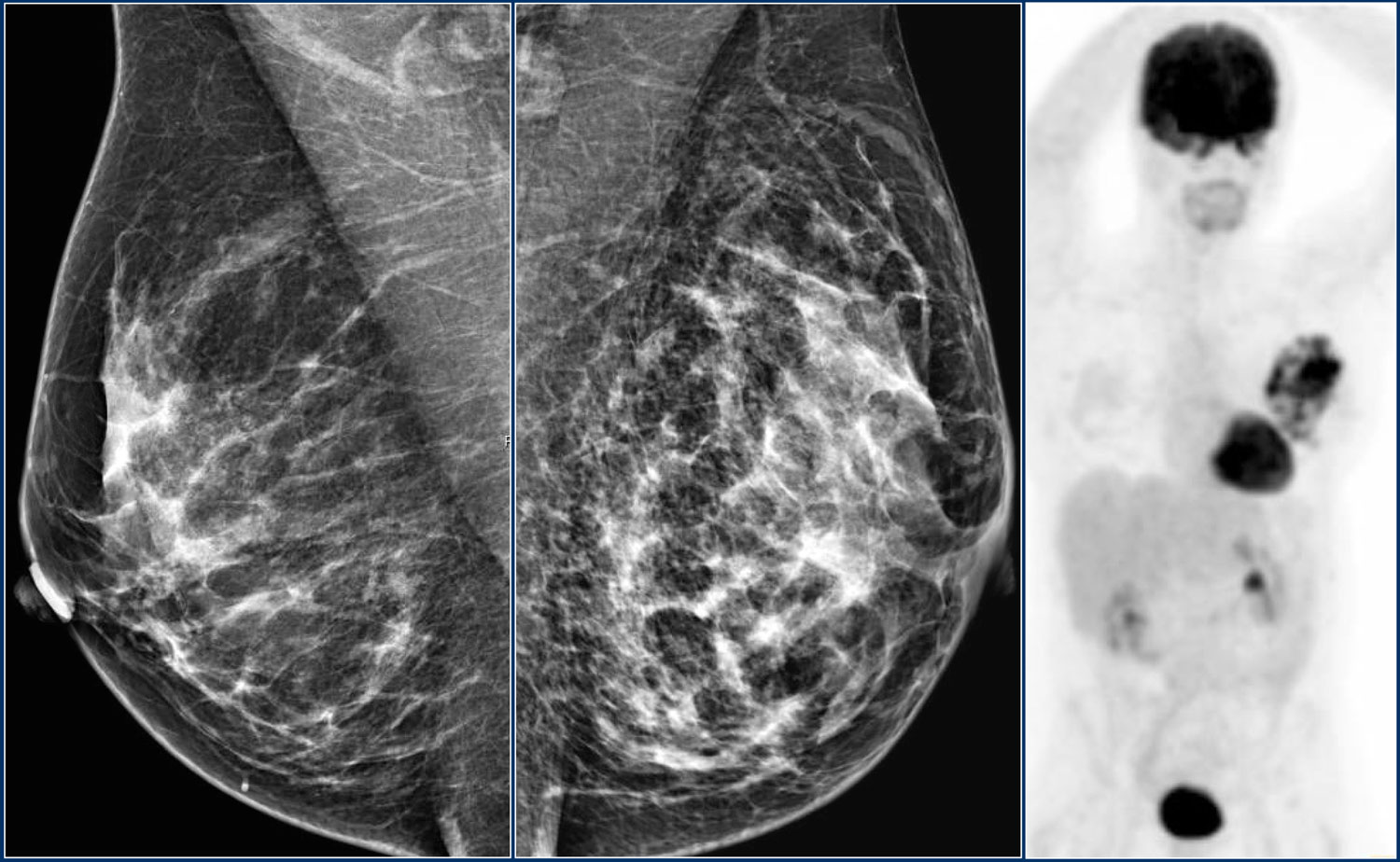
Что значит иметь плотную грудь?
Что такое плотность груди?
Плотность груди отражает количество фиброзной и железистой ткани в груди женщины по сравнению с количеством жировой ткани в груди, как видно на маммограмме.
В отчете о маммографии плотность груди относится к одной из следующих четырех категорий —
.- Грудь почти полностью жирная (около 10% женщин).
- Несколько участков плотной ткани разбросаны по груди (около 40% женщин).
- Грудь равномерно плотная на всем протяжении (около 40% женщин).
- Грудь очень плотная (около 10% женщин).
Считается, что у женщин первых двух категорий грудь малоплотная, не плотная или жирная. Считается, что у женщин, относящихся ко вторым двум категориям, высокая или плотная грудь. Около половины женщин в возрасте 40 лет и старше имеют плотную грудь.
Почему важна плотность груди?
Изображение слева показывает большую часть жирной груди, а изображение справа показывает плотную грудь.
CloseНа изображении слева показана в основном жирная грудь, а на изображении справа — плотная грудь.
Риск рака молочной железы
Женщины с плотной грудью имеют более высокий шанс заболеть раком груди. Чем плотнее ваша грудь, тем выше риск. Ученые не знают наверняка, почему это правда.
Пациенты с раком груди с плотной грудью на , а не на чаще умирают от рака, чем пациенты с неплотной (жирной) грудью.
Маммограммы
Плотная ткань может скрывать рак. Фиброзная и железистая ткань на маммограмме выглядит белой. Возможная опухоль тоже. Поскольку на маммограмме трудно отличить опухоль от плотной ткани груди, небольшую опухоль можно не заметить.
У кого больше шансов иметь плотную грудь?
Плотность вашей груди может меняться со временем. Как правило, у вас больше шансов получить плотную грудь, если вы —
- Моложе.
- Беременны или кормите грудью.
- Пройдите заместительную гормональную терапию.
- Имеют меньшую массу тела.
Что делать, если у меня плотная грудь?
Поговорите со своим врачом о вашем личном риске заболеть раком груди. Плотная грудь — лишь один из нескольких факторов риска рака груди. Ваш врач также будет думать о других факторах, таких как ваш возраст и семейный анамнез рака.
Различные тесты могут помочь обнаружить некоторые виды рака, которые не отображаются на маммограмме.Но эти тесты, скорее всего, дадут ложноположительных результатов (тест считается ненормальным, но на самом деле у вас нет рака). Ложноположительные результаты теста часто приводят к ненужным тестам, таким как биопсия. Кроме того, возможно, вам придется заплатить за эти тесты.
Ваш врач может предложить один из этих тестов —
- УЗИ груди. Аппарат, который использует звуковые волны для создания подробных изображений, называемых сонограммами, областей внутри груди.
- Магнитно-резонансная томография (МРТ). Разновидность сканирования тела, в которой используется магнит, подключенный к компьютеру. МРТ позволяет детально снимать участки внутри груди.
Поговорите со своим врачом о том, как часто вам следует проходить скрининг на рак груди и какие тесты рекомендует ваш врач.
Дополнительная информация
Что это такое и связано ли это с раком груди?
Рассеянная фиброгландулярная ткань молочной железы — доброкачественное или доброкачественное заболевание, при котором одна или обе груди могут ощущаться бугорками.Если кисты развиваются, это может быть болезненным.
Женская грудь содержит фиброзную и жировую ткань. Пропорции у разных людей различаются.
Женщины с высокой долей волокнистой соединительной и железистой ткани по сравнению с жировой тканью имеют грудь высокой плотности. У людей с высокой долей жировой ткани грудь низкой плотности.
Грудь с разбросанной фиброзно-желтой тканью содержит как фиброзную, так и жировую ткань. По данным Национального института рака, около 40% женщин имеют этот тип ткани груди.
Люди могут замечать опухоль и дискомфорт в определенные периоды жизни и во время месячного цикла, например, во время менструации. С возрастом доля жировой ткани может увеличиваться. Гормональные колебания со временем могут повлиять на плотность груди.
Хотя большинство уплотнений в груди не являются злокачественными, любой, кто обнаруживает уплотнение, должен обратиться к врачу для дальнейшего обследования, чтобы исключить вероятность рака.
Грудь различается по плотности у разных людей и с течением времени.
Маммограмма может дать информацию о плотности и составе груди.
Ткани груди могут быть:
Ткани низкой плотности : Грудь содержит в основном жировую ткань. По данным Управления по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA), около 10% женщин в Соединенных Штатах имеют такую ткань груди. Исследования показывают, что женщины, чья грудь в основном состоит из жировой ткани, имеют более низкий риск рака груди, чем женщины с другими типами.
Плотная или очень плотная ткань : Ткань груди плотная на всем протяжении. FDA также заявляет, что около 50% женщин в США.С. имеют плотную грудь. Люди с плотной грудью могут иметь более высокий риск развития рака груди, чем люди с низкой плотностью груди.
Рассеянная фиброгландулярная ткань : Грудь сочетает в себе жировую и рассеянную фиброгландулярную ткань. Статистика FDA показывает, что 40% женщин в США имеют такую ткань. Женщины с этим типом ткани имеют более высокий риск рака груди, чем женщины с низкой плотностью груди, но более низкий риск по сравнению с женщинами с высокой плотностью груди.
Особенности рассеянной фиброгландулярной ткани груди могут включать:
- уплотнения в груди
- кисты, которые представляют собой заполненные жидкостью круглые или овальные мешочки
- фиброз или выступающую рубцовидную фиброзную ткань
- чрезмерный рост клеток в груди. слизистая оболочка молочных протоков или производящих молоко тканей
- увеличенные доли груди или аденоз
Знание того, как грудь ощущается и как она изменяется в течение месячного цикла, может помочь человеку узнать, происходят ли необычные изменения и когда обращаться за медицинской помощью совет.
Если одни и те же изменения происходят каждый месяц, они вряд ли станут поводом для беспокойства. Любой, кто замечает новую шишку или изменение, которое не меняется в течение месяца, должен подумать о том, чтобы обратиться к врачу для осмотра.
Также стоит помнить, что уплотнения в груди часто возникают не из-за рака груди.
Есть много типов уплотнений в груди, включая кисты и фиброаденомы. Узнайте больше о том, как распознать потом и что делать, если вы его найдете.
Причины, по которым у одних людей наблюдается рассеяние фиброгландулярной ткани молочной железы, а у других, не остаются неясными.
Одним из факторов может быть наличие и колебания репродуктивных гормонов, таких как эстроген. Повышение и понижение уровня эстрогена во время менструального цикла может привести к изменениям груди и дискомфорту.
Нежные, опухшие и болезненные уплотнения могут появиться перед менструацией, но обычно исчезают после менструации.
Люди с семейным анамнезом плотной ткани молочной железы или рассеянной фиброгландулярной ткани молочной железы более подвержены этому заболеванию.
С возрастом грудь становится менее плотной.В результате изменится соотношение жира и фиброзной ткани, и фиброгландулярной ткани груди станет меньше.
Маммография может помочь диагностировать рассеянную фиброгландулярную ткань груди. Маммограмма покажет, присутствуют ли какие-либо уплотнения, но не может показать, что это за опухоль.
Только биопсия может определить, является ли опухоль злокачественной.
Во время биопсии врач берет ткань или жидкость из груди для исследования в лаборатории. Это покажет, является ли опухоль злокачественной, кистой или другим доброкачественным образованием.
Если биопсия выявляет рак, врач порекомендует дальнейшее лечение.
Что включает в себя биопсия груди? Узнайте больше здесь. Лекарства OTC могут помочь облегчить любую боль, вызванную разбросанной фиброгландулярной тканью груди.
Рассеивание фиброгландулярной ткани молочной железы не является заболеванием и не требует лечения.
При возникновении боли можно облегчить симптомы безрецептурными лекарствами, такими как ибупрофен.
Те, кто курят или потребляют большое количество кофеина, могут обнаружить, что комки уменьшаются, когда они бросают или сокращают потребление.
Наличие фиброгландулярной ткани груди не увеличивает риск развития рака груди, но может затруднить обнаружение изменений.
Американский колледж врачей рекомендует женщинам начинать говорить со своим врачом о скрининге с 40 лет. Они также рекомендуют людям со средним риском рака груди проходить маммографию каждые 2 года в возрасте от 50 до 74 лет.
Другие организации, например, Американское онкологическое общество, имеют другие руководящие принципы.
Врач порекомендует индивидуальный план обследования в зависимости от его обстоятельств.
Рассеянная фиброгландулярная ткань груди — обычное явление, которое может вызывать болезненные или опухшие груди в определенные периоды менструального цикла.
Это не рак и обычно не вызывает проблем со здоровьем, но наличие шишек в груди может усилить беспокойство по поводу рака.
Человек, знакомый со своей грудью и тем, как она меняется с течением времени, сможет заметить любые изменения, которые могут потребовать внимания.
Следование рекомендациям врача по скринингу может увеличить шансы на раннюю диагностику и быстрое лечение в случае возникновения проблемы.
Q:
У меня опухшая грудь с кистами. Могут ли они стать злокачественными?
A:
Важно понимать, что определение любого вида риска рака варьируется от человека к человеку. Если вы заметили какие-либо изменения по сравнению с исходной грудью, стоит поговорить со своим врачом и получить дополнительную информацию или провести подробный осмотр.
Большинство врачей и текущие руководства рекомендуют мужчинам и женщинам развивать «осведомленность о груди», чтобы они могли обнаруживать любые изменения как можно раньше. В целом люди будут иметь благоприятные перспективы, если врач диагностирует и лечит рак на ранней стадии.
Кристина Чун, MPH Ответы отражают мнение наших медицинских экспертов. Весь контент носит исключительно информационный характер и не может рассматриваться как медицинский совет.Мы выбрали связанные элементы, основываясь на качестве продуктов, и перечислили плюсы и минусы каждого, чтобы помочь вам определить, какой из них лучше всего подойдет вам.Мы сотрудничаем с некоторыми компаниями, которые продают эти продукты, что означает, что Healthline UK и наши партнеры могут получать часть доходов, если вы совершите покупку, используя ссылку (ссылки) выше.
Что это такое и связано ли это с раком груди?
Рассеянная фиброгландулярная ткань молочной железы — доброкачественное или доброкачественное заболевание, при котором одна или обе груди могут ощущаться бугорками. Если кисты развиваются, это может быть болезненным.
Женская грудь содержит фиброзную и жировую ткань. Пропорции у разных людей различаются.
Женщины с высокой долей волокнистой соединительной и железистой ткани по сравнению с жировой тканью имеют грудь высокой плотности. У людей с высокой долей жировой ткани грудь низкой плотности.
Грудь с разбросанной фиброзно-желтой тканью содержит как фиброзную, так и жировую ткань. По данным Национального института рака, около 40% женщин имеют этот тип ткани груди.
Люди могут замечать опухоль и дискомфорт в определенные периоды жизни и во время месячного цикла, например, во время менструации.С возрастом доля жировой ткани может увеличиваться. Гормональные колебания со временем могут повлиять на плотность груди.
Хотя большинство уплотнений в груди не являются злокачественными, любой, кто обнаруживает уплотнение, должен обратиться к врачу для дальнейшего обследования, чтобы исключить вероятность рака.
Грудь различается по плотности у разных людей и с течением времени.
Маммограмма может дать информацию о плотности и составе груди.
Ткани груди могут быть:
Ткани низкой плотности : Грудь содержит в основном жировую ткань.По данным Управления по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA), около 10% женщин в Соединенных Штатах имеют такую ткань груди. Исследования показывают, что женщины, чья грудь в основном состоит из жировой ткани, имеют более низкий риск рака груди, чем женщины с другими типами.
Плотная или очень плотная ткань : Ткань груди плотная на всем протяжении. FDA также заявляет, что около 50% женщин в США имеют плотную грудь. Люди с плотной грудью могут иметь более высокий риск развития рака груди, чем люди с низкой плотностью груди.
Рассеянная фиброгландулярная ткань : Грудь сочетает в себе жировую и рассеянную фиброгландулярную ткань. Статистика FDA показывает, что 40% женщин в США имеют такую ткань. Женщины с этим типом ткани имеют более высокий риск рака груди, чем женщины с низкой плотностью груди, но более низкий риск по сравнению с женщинами с высокой плотностью груди.
Особенности рассеянной фиброгландулярной ткани груди могут включать:
- уплотнения в груди
- кисты, которые представляют собой заполненные жидкостью круглые или овальные мешочки
- фиброз или выступающую рубцовидную фиброзную ткань
- чрезмерный рост клеток в груди. слизистая оболочка молочных протоков или производящих молоко тканей
- увеличенные доли груди или аденоз
Знание того, как грудь ощущается и как она изменяется в течение месячного цикла, может помочь человеку узнать, происходят ли необычные изменения и когда обращаться за медицинской помощью совет.
Если одни и те же изменения происходят каждый месяц, они вряд ли станут поводом для беспокойства. Любой, кто замечает новую шишку или изменение, которое не меняется в течение месяца, должен подумать о том, чтобы обратиться к врачу для осмотра.
Также стоит помнить, что уплотнения в груди часто возникают не из-за рака груди.
Есть много типов уплотнений в груди, включая кисты и фиброаденомы. Узнайте больше о том, как распознать потом и что делать, если вы его найдете.
Причины, по которым у одних людей наблюдается рассеяние фиброгландулярной ткани молочной железы, а у других, не остаются неясными.
Одним из факторов может быть наличие и колебания репродуктивных гормонов, таких как эстроген. Повышение и понижение уровня эстрогена во время менструального цикла может привести к изменениям груди и дискомфорту.
Нежные, опухшие и болезненные уплотнения могут появиться перед менструацией, но обычно исчезают после менструации.
Люди с семейным анамнезом плотной ткани молочной железы или рассеянной фиброгландулярной ткани молочной железы более подвержены этому заболеванию.
С возрастом грудь становится менее плотной.В результате изменится соотношение жира и фиброзной ткани, и фиброгландулярной ткани груди станет меньше.
Маммография может помочь диагностировать рассеянную фиброгландулярную ткань груди. Маммограмма покажет, присутствуют ли какие-либо уплотнения, но не может показать, что это за опухоль.
Только биопсия может определить, является ли опухоль злокачественной.
Во время биопсии врач берет ткань или жидкость из груди для исследования в лаборатории. Это покажет, является ли опухоль злокачественной, кистой или другим доброкачественным образованием.
Если биопсия выявляет рак, врач порекомендует дальнейшее лечение.
Что включает в себя биопсия груди? Узнайте больше здесь. Лекарства OTC могут помочь облегчить любую боль, вызванную разбросанной фиброгландулярной тканью груди.
Рассеивание фиброгландулярной ткани молочной железы не является заболеванием и не требует лечения.
При возникновении боли можно облегчить симптомы безрецептурными лекарствами, такими как ибупрофен.
Те, кто курят или потребляют большое количество кофеина, могут обнаружить, что комки уменьшаются, когда они бросают или сокращают потребление.
Наличие фиброгландулярной ткани груди не увеличивает риск развития рака груди, но может затруднить обнаружение изменений.
Американский колледж врачей рекомендует женщинам начинать говорить со своим врачом о скрининге с 40 лет. Они также рекомендуют людям со средним риском рака груди проходить маммографию каждые 2 года в возрасте от 50 до 74 лет.
Другие организации, например, Американское онкологическое общество, имеют другие руководящие принципы.
Врач порекомендует индивидуальный план обследования в зависимости от его обстоятельств.
Рассеянная фиброгландулярная ткань груди — обычное явление, которое может вызывать болезненные или опухшие груди в определенные периоды менструального цикла.
Это не рак и обычно не вызывает проблем со здоровьем, но наличие шишек в груди может усилить беспокойство по поводу рака.
Человек, знакомый со своей грудью и тем, как она меняется с течением времени, сможет заметить любые изменения, которые могут потребовать внимания.
Следование рекомендациям врача по скринингу может увеличить шансы на раннюю диагностику и быстрое лечение в случае возникновения проблемы.
Q:
У меня опухшая грудь с кистами. Могут ли они стать злокачественными?
A:
Важно понимать, что определение любого вида риска рака варьируется от человека к человеку. Если вы заметили какие-либо изменения по сравнению с исходной грудью, стоит поговорить со своим врачом и получить дополнительную информацию или провести подробный осмотр.
Большинство врачей и текущие руководства рекомендуют мужчинам и женщинам развивать «осведомленность о груди», чтобы они могли обнаруживать любые изменения как можно раньше. В целом люди будут иметь благоприятные перспективы, если врач диагностирует и лечит рак на ранней стадии.
Кристина Чун, MPH Ответы отражают мнение наших медицинских экспертов. Весь контент носит исключительно информационный характер и не может рассматриваться как медицинский совет.Мы выбрали связанные элементы, основываясь на качестве продуктов, и перечислили плюсы и минусы каждого, чтобы помочь вам определить, какой из них лучше всего подойдет вам.Мы сотрудничаем с некоторыми компаниями, которые продают эти продукты, что означает, что Healthline UK и наши партнеры могут получать часть доходов, если вы совершите покупку, используя ссылку (ссылки) выше.
Что это такое и связано ли это с раком груди?
Рассеянная фиброгландулярная ткань молочной железы — доброкачественное или доброкачественное заболевание, при котором одна или обе груди могут ощущаться бугорками. Если кисты развиваются, это может быть болезненным.
Женская грудь содержит фиброзную и жировую ткань. Пропорции у разных людей различаются.
Женщины с высокой долей волокнистой соединительной и железистой ткани по сравнению с жировой тканью имеют грудь высокой плотности. У людей с высокой долей жировой ткани грудь низкой плотности.
Грудь с разбросанной фиброзно-желтой тканью содержит как фиброзную, так и жировую ткань. По данным Национального института рака, около 40% женщин имеют этот тип ткани груди.
Люди могут замечать опухоль и дискомфорт в определенные периоды жизни и во время месячного цикла, например, во время менструации.С возрастом доля жировой ткани может увеличиваться. Гормональные колебания со временем могут повлиять на плотность груди.
Хотя большинство уплотнений в груди не являются злокачественными, любой, кто обнаруживает уплотнение, должен обратиться к врачу для дальнейшего обследования, чтобы исключить вероятность рака.
Грудь различается по плотности у разных людей и с течением времени.
Маммограмма может дать информацию о плотности и составе груди.
Ткани груди могут быть:
Ткани низкой плотности : Грудь содержит в основном жировую ткань.По данным Управления по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA), около 10% женщин в Соединенных Штатах имеют такую ткань груди. Исследования показывают, что женщины, чья грудь в основном состоит из жировой ткани, имеют более низкий риск рака груди, чем женщины с другими типами.
Плотная или очень плотная ткань : Ткань груди плотная на всем протяжении. FDA также заявляет, что около 50% женщин в США имеют плотную грудь. Люди с плотной грудью могут иметь более высокий риск развития рака груди, чем люди с низкой плотностью груди.
Рассеянная фиброгландулярная ткань : Грудь сочетает в себе жировую и рассеянную фиброгландулярную ткань. Статистика FDA показывает, что 40% женщин в США имеют такую ткань. Женщины с этим типом ткани имеют более высокий риск рака груди, чем женщины с низкой плотностью груди, но более низкий риск по сравнению с женщинами с высокой плотностью груди.
Особенности рассеянной фиброгландулярной ткани груди могут включать:
- уплотнения в груди
- кисты, которые представляют собой заполненные жидкостью круглые или овальные мешочки
- фиброз или выступающую рубцовидную фиброзную ткань
- чрезмерный рост клеток в груди. слизистая оболочка молочных протоков или производящих молоко тканей
- увеличенные доли груди или аденоз
Знание того, как грудь ощущается и как она изменяется в течение месячного цикла, может помочь человеку узнать, происходят ли необычные изменения и когда обращаться за медицинской помощью совет.
Если одни и те же изменения происходят каждый месяц, они вряд ли станут поводом для беспокойства. Любой, кто замечает новую шишку или изменение, которое не меняется в течение месяца, должен подумать о том, чтобы обратиться к врачу для осмотра.
Также стоит помнить, что уплотнения в груди часто возникают не из-за рака груди.
Есть много типов уплотнений в груди, включая кисты и фиброаденомы. Узнайте больше о том, как распознать потом и что делать, если вы его найдете.
Причины, по которым у одних людей наблюдается рассеяние фиброгландулярной ткани молочной железы, а у других, не остаются неясными.
Одним из факторов может быть наличие и колебания репродуктивных гормонов, таких как эстроген. Повышение и понижение уровня эстрогена во время менструального цикла может привести к изменениям груди и дискомфорту.
Нежные, опухшие и болезненные уплотнения могут появиться перед менструацией, но обычно исчезают после менструации.
Люди с семейным анамнезом плотной ткани молочной железы или рассеянной фиброгландулярной ткани молочной железы более подвержены этому заболеванию.
С возрастом грудь становится менее плотной.В результате изменится соотношение жира и фиброзной ткани, и фиброгландулярной ткани груди станет меньше.
Маммография может помочь диагностировать рассеянную фиброгландулярную ткань груди. Маммограмма покажет, присутствуют ли какие-либо уплотнения, но не может показать, что это за опухоль.
Только биопсия может определить, является ли опухоль злокачественной.
Во время биопсии врач берет ткань или жидкость из груди для исследования в лаборатории. Это покажет, является ли опухоль злокачественной, кистой или другим доброкачественным образованием.
Если биопсия выявляет рак, врач порекомендует дальнейшее лечение.
Что включает в себя биопсия груди? Узнайте больше здесь. Лекарства OTC могут помочь облегчить любую боль, вызванную разбросанной фиброгландулярной тканью груди.
Рассеивание фиброгландулярной ткани молочной железы не является заболеванием и не требует лечения.
При возникновении боли можно облегчить симптомы безрецептурными лекарствами, такими как ибупрофен.
Те, кто курят или потребляют большое количество кофеина, могут обнаружить, что комки уменьшаются, когда они бросают или сокращают потребление.
Наличие фиброгландулярной ткани груди не увеличивает риск развития рака груди, но может затруднить обнаружение изменений.
Американский колледж врачей рекомендует женщинам начинать говорить со своим врачом о скрининге с 40 лет. Они также рекомендуют людям со средним риском рака груди проходить маммографию каждые 2 года в возрасте от 50 до 74 лет.
Другие организации, например, Американское онкологическое общество, имеют другие руководящие принципы.
Врач порекомендует индивидуальный план обследования в зависимости от его обстоятельств.
Рассеянная фиброгландулярная ткань груди — обычное явление, которое может вызывать болезненные или опухшие груди в определенные периоды менструального цикла.
Это не рак и обычно не вызывает проблем со здоровьем, но наличие шишек в груди может усилить беспокойство по поводу рака.
Человек, знакомый со своей грудью и тем, как она меняется с течением времени, сможет заметить любые изменения, которые могут потребовать внимания.
Следование рекомендациям врача по скринингу может увеличить шансы на раннюю диагностику и быстрое лечение в случае возникновения проблемы.
Q:
У меня опухшая грудь с кистами. Могут ли они стать злокачественными?
A:
Важно понимать, что определение любого вида риска рака варьируется от человека к человеку. Если вы заметили какие-либо изменения по сравнению с исходной грудью, стоит поговорить со своим врачом и получить дополнительную информацию или провести подробный осмотр.
Большинство врачей и текущие руководства рекомендуют мужчинам и женщинам развивать «осведомленность о груди», чтобы они могли обнаруживать любые изменения как можно раньше. В целом люди будут иметь благоприятные перспективы, если врач диагностирует и лечит рак на ранней стадии.
Кристина Чун, MPH Ответы отражают мнение наших медицинских экспертов. Весь контент носит исключительно информационный характер и не может рассматриваться как медицинский совет.Мы выбрали связанные элементы, основываясь на качестве продуктов, и перечислили плюсы и минусы каждого, чтобы помочь вам определить, какой из них лучше всего подойдет вам.Мы сотрудничаем с некоторыми компаниями, которые продают эти продукты, что означает, что Healthline UK и наши партнеры могут получать часть доходов, если вы совершите покупку, используя ссылку (ссылки) выше.
Что это такое и связано ли это с раком груди?
Рассеянная фиброгландулярная ткань молочной железы — доброкачественное или доброкачественное заболевание, при котором одна или обе груди могут ощущаться бугорками. Если кисты развиваются, это может быть болезненным.
Женская грудь содержит фиброзную и жировую ткань. Пропорции у разных людей различаются.
Женщины с высокой долей волокнистой соединительной и железистой ткани по сравнению с жировой тканью имеют грудь высокой плотности. У людей с высокой долей жировой ткани грудь низкой плотности.
Грудь с разбросанной фиброзно-желтой тканью содержит как фиброзную, так и жировую ткань. По данным Национального института рака, около 40% женщин имеют этот тип ткани груди.
Люди могут замечать опухоль и дискомфорт в определенные периоды жизни и во время месячного цикла, например, во время менструации.С возрастом доля жировой ткани может увеличиваться. Гормональные колебания со временем могут повлиять на плотность груди.
Хотя большинство уплотнений в груди не являются злокачественными, любой, кто обнаруживает уплотнение, должен обратиться к врачу для дальнейшего обследования, чтобы исключить вероятность рака.
Грудь различается по плотности у разных людей и с течением времени.
Маммограмма может дать информацию о плотности и составе груди.
Ткани груди могут быть:
Ткани низкой плотности : Грудь содержит в основном жировую ткань.По данным Управления по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA), около 10% женщин в Соединенных Штатах имеют такую ткань груди. Исследования показывают, что женщины, чья грудь в основном состоит из жировой ткани, имеют более низкий риск рака груди, чем женщины с другими типами.
Плотная или очень плотная ткань : Ткань груди плотная на всем протяжении. FDA также заявляет, что около 50% женщин в США имеют плотную грудь. Люди с плотной грудью могут иметь более высокий риск развития рака груди, чем люди с низкой плотностью груди.
Рассеянная фиброгландулярная ткань : Грудь сочетает в себе жировую и рассеянную фиброгландулярную ткань. Статистика FDA показывает, что 40% женщин в США имеют такую ткань. Женщины с этим типом ткани имеют более высокий риск рака груди, чем женщины с низкой плотностью груди, но более низкий риск по сравнению с женщинами с высокой плотностью груди.
Особенности рассеянной фиброгландулярной ткани груди могут включать:
- уплотнения в груди
- кисты, которые представляют собой заполненные жидкостью круглые или овальные мешочки
- фиброз или выступающую рубцовидную фиброзную ткань
- чрезмерный рост клеток в груди. слизистая оболочка молочных протоков или производящих молоко тканей
- увеличенные доли груди или аденоз
Знание того, как грудь ощущается и как она изменяется в течение месячного цикла, может помочь человеку узнать, происходят ли необычные изменения и когда обращаться за медицинской помощью совет.
Если одни и те же изменения происходят каждый месяц, они вряд ли станут поводом для беспокойства. Любой, кто замечает новую шишку или изменение, которое не меняется в течение месяца, должен подумать о том, чтобы обратиться к врачу для осмотра.
Также стоит помнить, что уплотнения в груди часто возникают не из-за рака груди.
Есть много типов уплотнений в груди, включая кисты и фиброаденомы. Узнайте больше о том, как распознать потом и что делать, если вы его найдете.
Причины, по которым у одних людей наблюдается рассеяние фиброгландулярной ткани молочной железы, а у других, не остаются неясными.
Одним из факторов может быть наличие и колебания репродуктивных гормонов, таких как эстроген. Повышение и понижение уровня эстрогена во время менструального цикла может привести к изменениям груди и дискомфорту.
Нежные, опухшие и болезненные уплотнения могут появиться перед менструацией, но обычно исчезают после менструации.
Люди с семейным анамнезом плотной ткани молочной железы или рассеянной фиброгландулярной ткани молочной железы более подвержены этому заболеванию.
С возрастом грудь становится менее плотной.В результате изменится соотношение жира и фиброзной ткани, и фиброгландулярной ткани груди станет меньше.
Маммография может помочь диагностировать рассеянную фиброгландулярную ткань груди. Маммограмма покажет, присутствуют ли какие-либо уплотнения, но не может показать, что это за опухоль.
Только биопсия может определить, является ли опухоль злокачественной.
Во время биопсии врач берет ткань или жидкость из груди для исследования в лаборатории. Это покажет, является ли опухоль злокачественной, кистой или другим доброкачественным образованием.
Если биопсия выявляет рак, врач порекомендует дальнейшее лечение.
Что включает в себя биопсия груди? Узнайте больше здесь. Лекарства OTC могут помочь облегчить любую боль, вызванную разбросанной фиброгландулярной тканью груди.
Рассеивание фиброгландулярной ткани молочной железы не является заболеванием и не требует лечения.
При возникновении боли можно облегчить симптомы безрецептурными лекарствами, такими как ибупрофен.
Те, кто курят или потребляют большое количество кофеина, могут обнаружить, что комки уменьшаются, когда они бросают или сокращают потребление.
Наличие фиброгландулярной ткани груди не увеличивает риск развития рака груди, но может затруднить обнаружение изменений.
Американский колледж врачей рекомендует женщинам начинать говорить со своим врачом о скрининге с 40 лет. Они также рекомендуют людям со средним риском рака груди проходить маммографию каждые 2 года в возрасте от 50 до 74 лет.
Другие организации, например, Американское онкологическое общество, имеют другие руководящие принципы.
Врач порекомендует индивидуальный план обследования в зависимости от его обстоятельств.
Рассеянная фиброгландулярная ткань груди — обычное явление, которое может вызывать болезненные или опухшие груди в определенные периоды менструального цикла.
Это не рак и обычно не вызывает проблем со здоровьем, но наличие шишек в груди может усилить беспокойство по поводу рака.
Человек, знакомый со своей грудью и тем, как она меняется с течением времени, сможет заметить любые изменения, которые могут потребовать внимания.
Следование рекомендациям врача по скринингу может увеличить шансы на раннюю диагностику и быстрое лечение в случае возникновения проблемы.
Q:
У меня опухшая грудь с кистами. Могут ли они стать злокачественными?
A:
Важно понимать, что определение любого вида риска рака варьируется от человека к человеку. Если вы заметили какие-либо изменения по сравнению с исходной грудью, стоит поговорить со своим врачом и получить дополнительную информацию или провести подробный осмотр.
Большинство врачей и текущие руководства рекомендуют мужчинам и женщинам развивать «осведомленность о груди», чтобы они могли обнаруживать любые изменения как можно раньше. В целом люди будут иметь благоприятные перспективы, если врач диагностирует и лечит рак на ранней стадии.
Кристина Чун, MPH Ответы отражают мнение наших медицинских экспертов. Весь контент носит исключительно информационный характер и не может рассматриваться как медицинский совет.Мы выбрали связанные элементы, основываясь на качестве продуктов, и перечислили плюсы и минусы каждого, чтобы помочь вам определить, какой из них лучше всего подойдет вам.Мы сотрудничаем с некоторыми компаниями, которые продают эти продукты, что означает, что Healthline UK и наши партнеры могут получать часть доходов, если вы совершите покупку, используя ссылку (ссылки) выше.
Фиброгландулярная плотность | Определение плотности груди и волокнистых тканей в центрах здоровья груди UH
Плотная ткань груди может затруднить выявление рака и подвергнуть женщин более высокому риску
Download Плотность груди: скрининг рака грудиГрудь состоит из смеси фиброзной, железистой и жировой ткани.Грудь женщины считается плотной, если у нее много фиброзной или железистой ткани, но мало жира. Плотность может уменьшаться с возрастом, но у большинства женщин это практически не меняется.
Плотность груди определяется рентгенологом, который читает маммограмму женщины. Маммограмма определяет четыре категории плотности. Радиолог относит каждую маммограмму к одной из категорий. Врач женщины должен определить, плотная ли у нее грудь, по тому месту, где она находится на шкале плотности.
Четыре уровня плотности груди:
- Почти полностью жирный
- Рассеянные участки фиброзно-желёзной плотности
- Неоднородная плотность
- Чрезвычайно плотный
Радиологи классифицируют плотность груди по 4-уровневой шкале плотности:
Почти полностью жирный
Рассеянные участки фиброгландулярной плотности
Неоднородно плотный
Чрезвычайно плотная
В U.С., Плотность груди распределяется следующим образом:
- 10 процентов женщин имеют почти полностью жирную грудь
- 10% имеют очень плотную грудь
- 80 процентов относятся к одной из двух средних категорий
Почему важна плотность груди?
Наличие плотной ткани груди может увеличить вероятность заболевания раком груди у женщины. Плотная грудь также затрудняет врачам выявление рака на маммограмме. Плотная ткань выглядит белой на маммограмме; уплотнения, как доброкачественные, так и злокачественные, также выглядят белыми.Таким образом, маммография может быть менее точной у женщин с плотной грудью.
Если маммограмма показывает, что у женщины неоднородно плотная или чрезвычайно плотная ткань груди, она получит письмо с уведомлением о том, что у нее плотная грудь. Затем она может поговорить со своим лечащим врачом о том, что это значит для нее и какие дополнительные тесты могут потребоваться, если таковые имеются.
Плотная грудь: ответы на часто задаваемые вопросы
Что такое плотная грудь?
Грудь содержит железистую, соединительную и жировую ткань.Плотность груди — это термин, который описывает относительное количество этих различных типов ткани груди, как видно на маммограмме. Плотная грудь имеет относительно большое количество железистой и волокнистой соединительной ткани и относительно небольшое количество жировой ткани груди.
Как узнать, плотная ли у меня грудь?
Только маммография может показать, если у женщины плотная грудь. Плотная ткань груди не может быть обнаружена при клиническом осмотре груди или при самообследовании груди. По этой причине плотную грудь иногда называют маммографически плотной грудью.
Насколько распространена плотная грудь?
Почти половина всех женщин в возрасте 40 лет и старше, сделавших маммографию, имеют плотную грудь. Плотность груди часто передается по наследству, но на нее могут влиять другие факторы. Факторы, связанные с более низкой плотностью груди, включают возраст, рождение детей и прием тамоксифена. Факторы, связанные с более высокой плотностью груди, включают использование заместительной гормональной терапии в постменопаузе и низкий индекс массы тела.
Как классифицируется плотность груди?
Врачи используют систему отчетов и данных визуализации груди, называемую BI-RADS, для группировки различных типов плотности груди.Эта система, разработанная Американским колледжем радиологии, помогает врачам интерпретировать результаты маммограммы и сообщать о них. Врачи, просматривающие маммограммы, называются радиологами. BI-RADS классифицирует плотность груди на четыре категории, а именно:
Если вам говорят, что у вас плотная грудь, это означает, что у вас либо «неоднородно плотная» (C), либо «чрезвычайно плотная» (D) грудь.
На этом изображении показаны четыре категории плотности груди.Грудь может быть почти полностью жирной (A), иметь отдельные участки плотной фиброгландулярной ткани (B), иметь много участков железистой и соединительной ткани (C) или быть чрезвычайно плотной (D). Грудь классифицируется как «плотная», если она попадает в категории неоднородно плотной (C) или чрезвычайно плотной (D).
Влияет ли плотная ткань груди на маммограмму женщины?
Маммограммы труднее читать женщинам с плотной грудью, чем женщинам с жирной грудью.Это связано с тем, что плотная ткань груди и некоторые аномальные изменения груди, такие как кальцификаты и опухоли, отображаются на маммограмме как белые участки. В результате маммография менее чувствительна у женщин с плотной грудью, то есть с большей вероятностью пропустить рак. Женщин с плотной грудью могут вызывать на контрольные обследования чаще, чем женщин с жирной грудью. Плотная грудь также повышает риск развития интервального рака груди у женщин. Это рак груди, который диагностируется в течение 12 месяцев после нормального результата маммографии.
Является ли плотная грудь фактором риска рака груди?
Да, женщины с плотной грудью имеют более высокий риск рака груди, чем женщины с жирной грудью, и этот риск увеличивается с увеличением плотности груди. Этот повышенный риск не связан с влиянием плотной груди на способность читать маммограмму.
Вероятность смерти от рака груди у больных раком груди с плотной грудью выше?
Нет. Исследования показали, что пациенты с раком груди с плотной грудью не более склонны к смерти от рака, чем пациенты с раком груди с жирной грудью, с учетом других факторов здоровья и характеристик опухоли.
Следует ли женщинам с плотной грудью проходить дополнительный скрининг на рак груди?
В некоторых штатах провайдеры маммографии обязаны сообщать женщинам, которым сделана маммограмма, плотность груди в целом или ее плотность. Многие штаты теперь требуют, чтобы женщины с плотной грудью были застрахованы на дополнительные визуализационные тесты. Карта США с информацией о законодательстве конкретного штата доступна на сайте DenseBreast-info.org.
Тем не менее, в соответствии с Заключительным заявлением о рекомендациях по скринингу рака груди, подготовленным Целевой группой профилактических служб США, ценность дополнительных или дополнительных скрининговых тестов, таких как УЗИ или МРТ, для женщин с плотной грудью еще не ясна.Текущие клинические испытания оценивают роль дополнительных методов визуализации у женщин с плотной грудью. Информационная служба рака NCI может рассказать вам о клинических испытаниях и предоставить индивидуальный поиск по клиническим испытаниям, чтобы помочь вам узнать больше о клинических испытаниях, связанных с плотностью груди и скринингом рака груди.
Недавнее исследование показало, что для женщин с плотной грудью стратегия скрининга, которая также учитывает факторы риска и защитные факторы женщины, может быть лучшим предиктором того, разовьется ли у женщины рак груди после нормальной маммографии и до ее следующей запланированной маммографии. .
Обсуждая с врачом ваш личный риск рака груди, имейте в виду, что:
Сводка PDQ® по профилактике рака молочной железыNCI включает раздел, в котором перечислены факторы риска и защитные факторы рака молочной железы.
Может быть полезно задать своему врачу следующие вопросы, чтобы рассмотреть ваш риск рака груди в контексте:
- Каковы результаты моей недавней маммографии?
- Рекомендуются ли другие дополнительные скрининговые или диагностические тесты?
- Каковы мои личные факторы риска рака груди? Защитные факторы при раке груди?
- Какие шаги я могу предпринять, чтобы снизить риск рака груди?
Что исследователи надеются узнать о взаимосвязи между плотностью груди и раком груди?
Некоторые важные вопросы включают:
- Могут ли визуализационные тесты, такие как трехмерная маммография (томосинтез груди), МРТ и УЗИ, помочь получить более четкую картину плотности груди?
- Существуют ли определенные узоры или участки плотной ткани груди, которые являются особенно «опасными»?
- Почему у некоторых женщин с плотной грудью развивается рак груди, а у других — нет? Можно ли определить биомаркеры, которые помогут предсказать, разовьется ли рак груди у женщины с плотной грудью?
- Связаны ли изменения плотности груди с течением времени с изменениями риска рака груди?
- Могут ли женщины снизить плотность груди и, возможно, риск рака груди, принимая лекарства или применяя местные агенты непосредственно на груди?
Подразделение профилактики рака (DCP) NCI поддерживает исследования в области скрининга рака и факторов риска, включая плотность груди, такие как это клиническое испытание для женщин с плотной грудью.
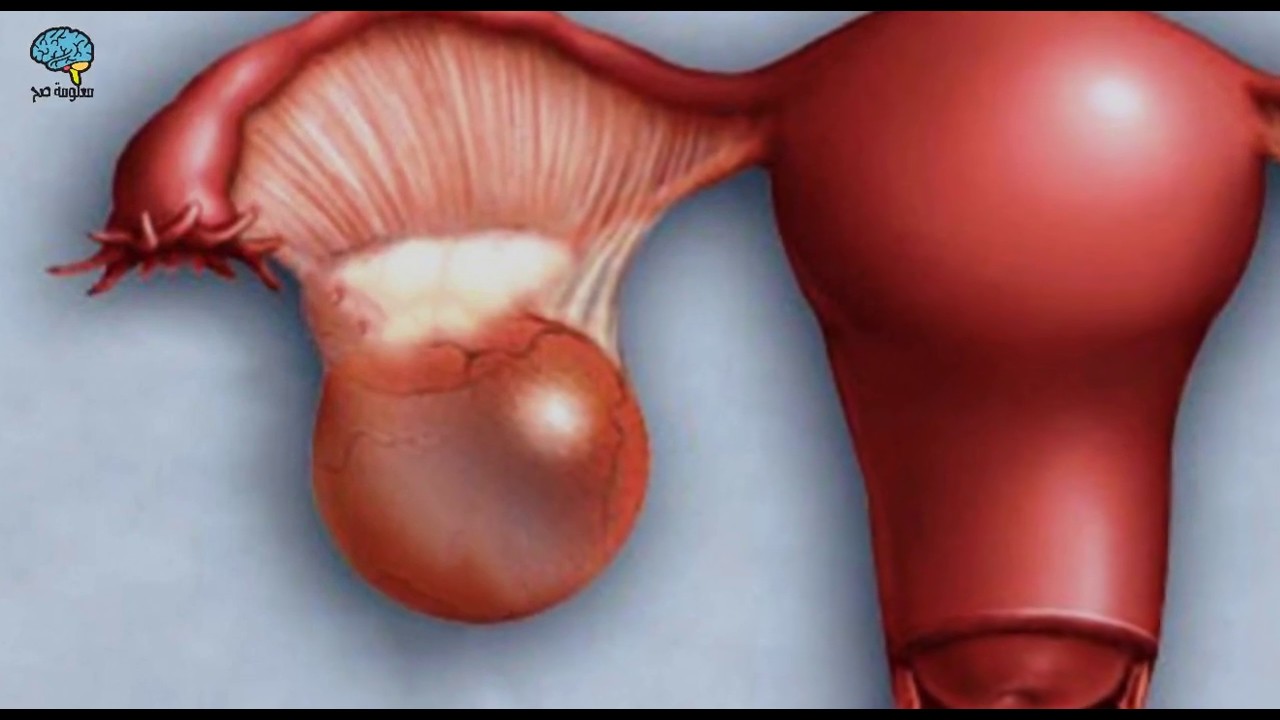 Обычно имеет тянущий и ноющий характер. Может отдавать в поясницу.
Обычно имеет тянущий и ноющий характер. Может отдавать в поясницу. Киста может образоваться также на фоне эндокринологических заболеваний, генетической предрасположенности (если такая проблема случалась у женщин вашего рода), при нарушении обменных процессов в организме. Также образоваться киста может при наличии воспалительных заболеваний малого таза. Возможно, вы слышали, что причина данного заболевания – переохлаждение. И это, действительно, может стать толчком для развития кисты.
Киста может образоваться также на фоне эндокринологических заболеваний, генетической предрасположенности (если такая проблема случалась у женщин вашего рода), при нарушении обменных процессов в организме. Также образоваться киста может при наличии воспалительных заболеваний малого таза. Возможно, вы слышали, что причина данного заболевания – переохлаждение. И это, действительно, может стать толчком для развития кисты. Киста может стать причиной бесплодия, а ее разрыв приводит к кровоизлиянию внутри брюшной полости. Среди осложнений встречаются: перекрут кисты, нагноение. Все перечисленные осложнения требуют немедленного хирургического вмешательства.
Киста может стать причиной бесплодия, а ее разрыв приводит к кровоизлиянию внутри брюшной полости. Среди осложнений встречаются: перекрут кисты, нагноение. Все перечисленные осложнения требуют немедленного хирургического вмешательства.  Как правило, кисты образуются у женщин до периода менопаузы. Но это не значит, что с наступлением менопаузы посещение гинеколога не требуется. Берегите женское здоровье, регулярно посещайте гинеколога и своевременно реагируйте на сигналы организма о возможных нарушениях.
Как правило, кисты образуются у женщин до периода менопаузы. Но это не значит, что с наступлением менопаузы посещение гинеколога не требуется. Берегите женское здоровье, регулярно посещайте гинеколога и своевременно реагируйте на сигналы организма о возможных нарушениях.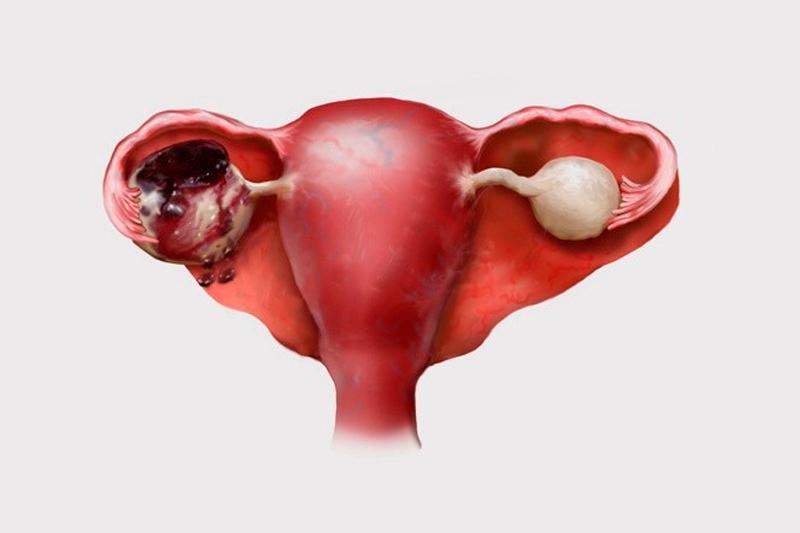 Хотя и здесь есть опасность, т.к. всегда существуют обстоятельства, которые могут спровоцировать разрыв и перекрут кисты, что потребует немедленного оперативного вмешательства.
Хотя и здесь есть опасность, т.к. всегда существуют обстоятельства, которые могут спровоцировать разрыв и перекрут кисты, что потребует немедленного оперативного вмешательства.
 Операция по удалению кисты яичника в АМК проводится на современном оборудовании специалистами высшей категории.
Операция по удалению кисты яичника в АМК проводится на современном оборудовании специалистами высшей категории. Но если киста яичника функциональная, которая возникает вследствие не произошедшей овуляции или накопления избыточной жидкости после овуляции, в большинстве случаев врачи не рекомендуют оперативное вмешательство. В период следующей менструации она может самостоятельно исчезнуть. Но стоит помнить, что любые новообразования, говорят об общем дисбалансе в организме.
Но если киста яичника функциональная, которая возникает вследствие не произошедшей овуляции или накопления избыточной жидкости после овуляции, в большинстве случаев врачи не рекомендуют оперативное вмешательство. В период следующей менструации она может самостоятельно исчезнуть. Но стоит помнить, что любые новообразования, говорят об общем дисбалансе в организме.

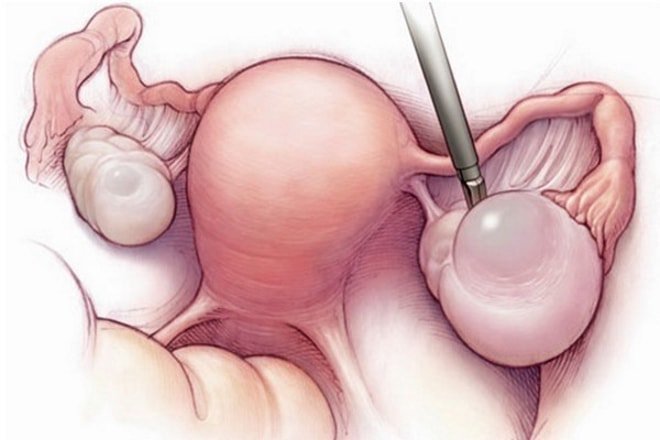
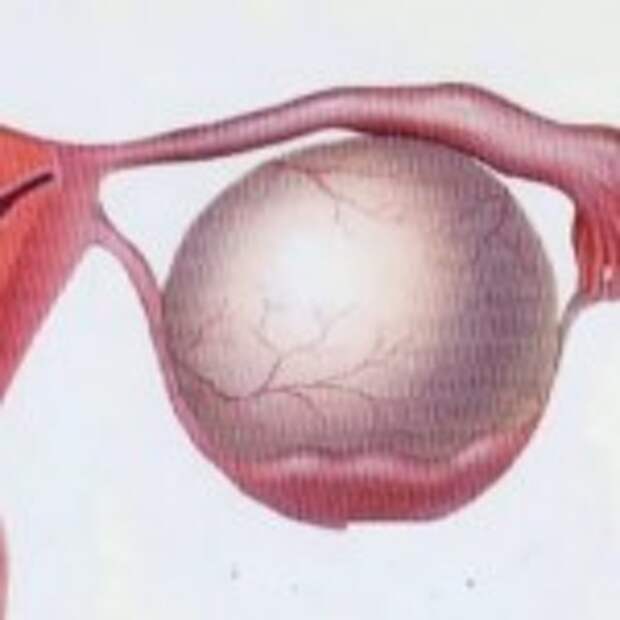
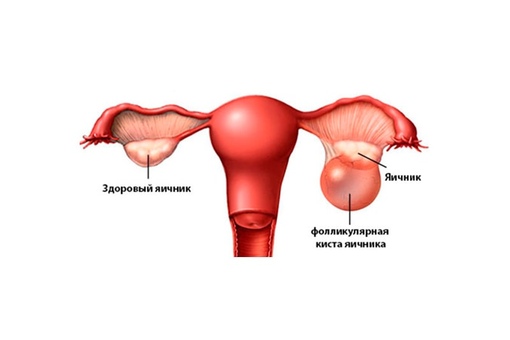

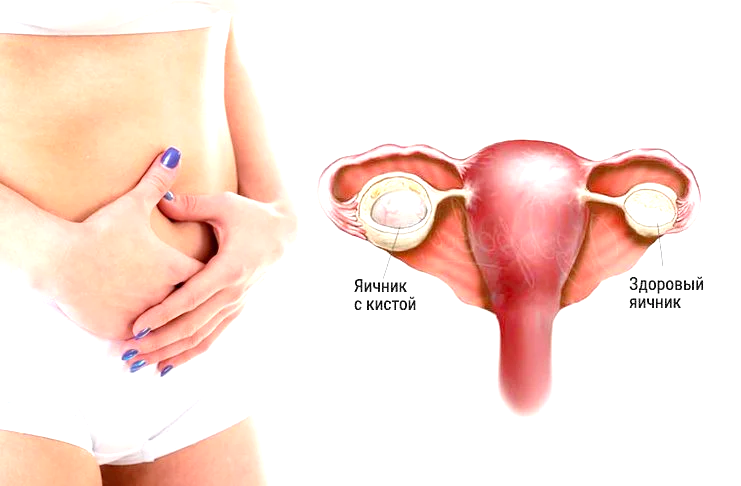 Эндометриоидная киста – это последствие эндометриоза – заболевания, при котором клетки внутреннего слоя стенки матки разрастаются.
Эндометриоидная киста – это последствие эндометриоза – заболевания, при котором клетки внутреннего слоя стенки матки разрастаются.
 Обычно она не перерождается в злокачественное образование, но может значительно расти в размерах, сдавливая при этом стенки мочевого пузыря и/или прямой кишки, нарушая тем самым работу этих органов.
Обычно она не перерождается в злокачественное образование, но может значительно расти в размерах, сдавливая при этом стенки мочевого пузыря и/или прямой кишки, нарушая тем самым работу этих органов.

 В отличие от опухолей киста не прорастает в органы, не дает метастаз и не производит гормоны. Функциональные кисты встречаются только у девочек и женщин репродуктивного возраста.
В отличие от опухолей киста не прорастает в органы, не дает метастаз и не производит гормоны. Функциональные кисты встречаются только у девочек и женщин репродуктивного возраста. Специалист осматривает полость малого таза, яичники и само образование, делая вывод о его размерах и структуре. УЗИ необходимо сделать несколько раз: если через 4-6 недель после первого УЗИ киста увеличилась, то врач предполагает опухоль и назначает дополнительные исследования.
Специалист осматривает полость малого таза, яичники и само образование, делая вывод о его размерах и структуре. УЗИ необходимо сделать несколько раз: если через 4-6 недель после первого УЗИ киста увеличилась, то врач предполагает опухоль и назначает дополнительные исследования.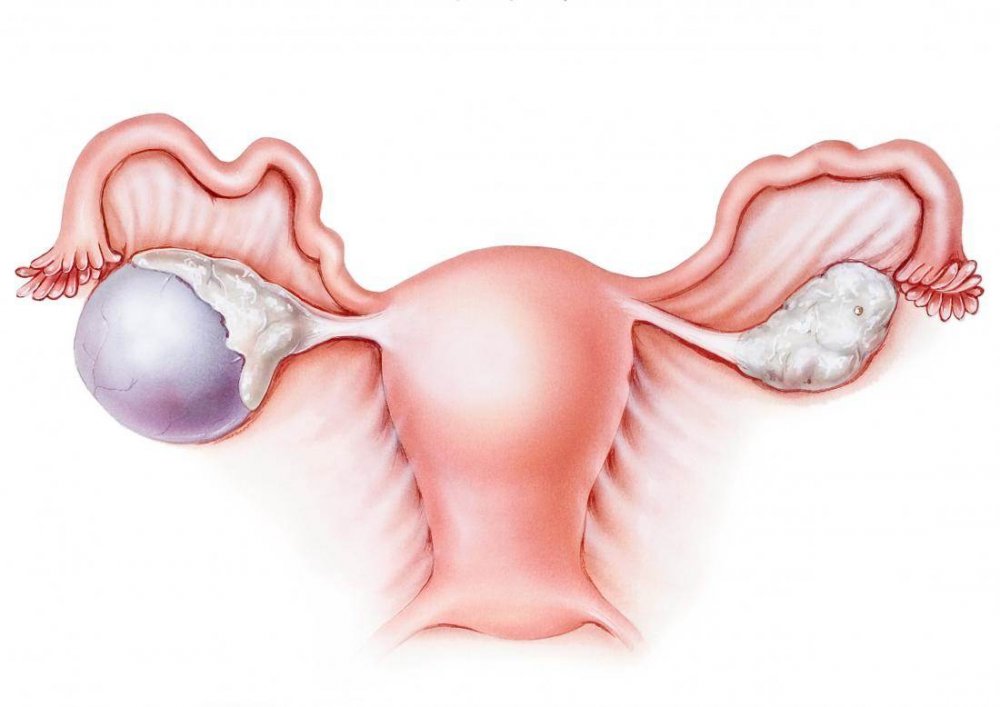

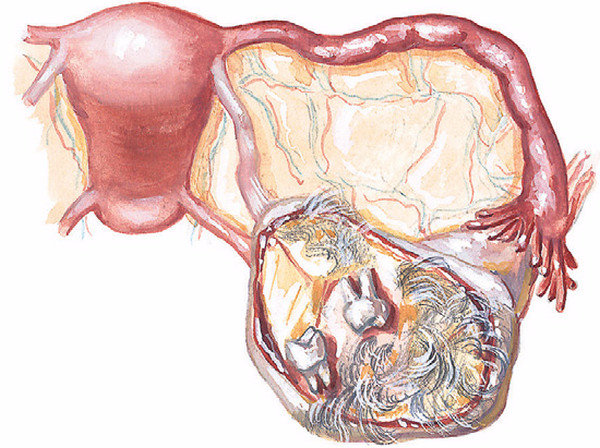 м.н., врач высшей категории
м.н., врач высшей категории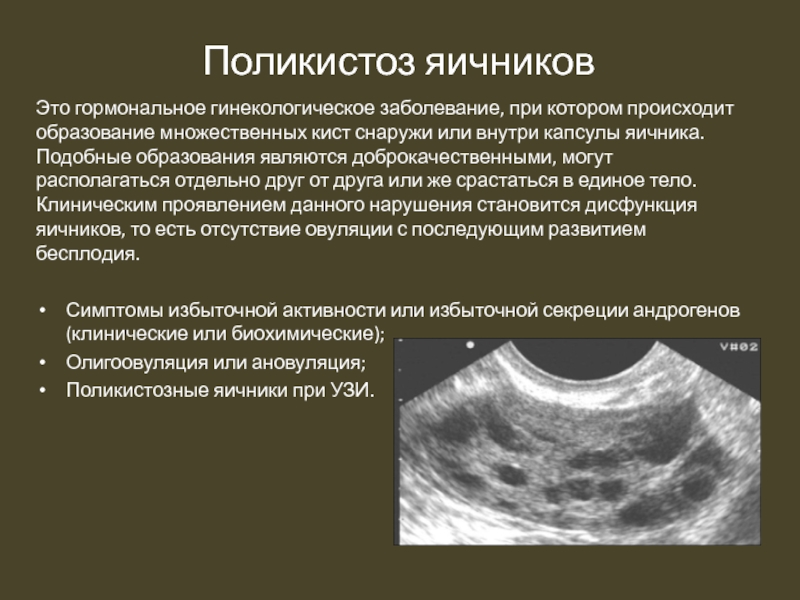 м.н.
м.н.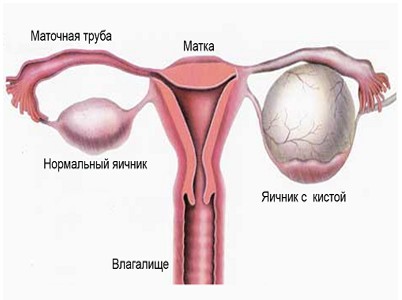 м.н.
м.н. Сенежская (ст. МЦК «Коптево»)
Сенежская (ст. МЦК «Коптево») Сенежская (ст. МЦК «Коптево»)
Сенежская (ст. МЦК «Коптево») «ВДНХ»)
«ВДНХ») м.н.
м.н. Расковой (м. «Белорусская»)
Расковой (м. «Белорусская») м.н., заведующая отделения ВРТ
м.н., заведующая отделения ВРТ м.н.
м.н. «Севастопольская»)
«Севастопольская») м.н.
м.н.

 Это может стать болезненным. Также есть вероятность рака, но это редко. С возрастом шансы возрастают.
Это может стать болезненным. Также есть вероятность рака, но это редко. С возрастом шансы возрастают. Выстелите блюдо салатными листьями или украсьте края кружками свежих огурцов. Аккуратно переложите скумбрию с фольги на блюдо. Украсьте рыбу веточками свежей зелени, кусочками маслин и дольками лимона или лайма.
Выстелите блюдо салатными листьями или украсьте края кружками свежих огурцов. Аккуратно переложите скумбрию с фольги на блюдо. Украсьте рыбу веточками свежей зелени, кусочками маслин и дольками лимона или лайма. Простые и вкусные рецепты приготовления отлично подходят для правильного питания, не требуют недоступных продуктов, по ним быстро и легко готовить. Если хочется уменьшить калорийность, то лучше запекать без обжарки овощей, жирных сливок, майонеза и сыра.
Простые и вкусные рецепты приготовления отлично подходят для правильного питания, не требуют недоступных продуктов, по ним быстро и легко готовить. Если хочется уменьшить калорийность, то лучше запекать без обжарки овощей, жирных сливок, майонеза и сыра. Смешать вместе горчицу, соус соевый, соль и любимые специи. На тушке сделать небольшие надрезы и поместить ее в маринад, оставить на один час. Решетку смазать маслом, выложить рыбу и вложить в разрезы кружки лимона. Готовить блюдо в духовке на решетке полчаса при 180 градусах.
Смешать вместе горчицу, соус соевый, соль и любимые специи. На тушке сделать небольшие надрезы и поместить ее в маринад, оставить на один час. Решетку смазать маслом, выложить рыбу и вложить в разрезы кружки лимона. Готовить блюдо в духовке на решетке полчаса при 180 градусах. Переложить скумбрию в стеклянную форму или на противень с фольгой, в брюшко можно добавить оставшийся апельсин, завернуть в фольгу. Духовку разогреть до 220 градусов, готовить блюдо 20 минут, затем открыть фольгу и оставить еще на 10 минут.
Переложить скумбрию в стеклянную форму или на противень с фольгой, в брюшко можно добавить оставшийся апельсин, завернуть в фольгу. Духовку разогреть до 220 градусов, готовить блюдо 20 минут, затем открыть фольгу и оставить еще на 10 минут. Готовить можно в стеклянной форме или противне, но предварительно надо застелить фольгой. Выложить и нафаршировать скумбрию овощной смесью, закрыть фольгой и запекать в духовке сорок минут на 180 градусах. За 5-10 минут до готовности приоткрыть фольгу и засыпать натертый сыр.
Готовить можно в стеклянной форме или противне, но предварительно надо застелить фольгой. Выложить и нафаршировать скумбрию овощной смесью, закрыть фольгой и запекать в духовке сорок минут на 180 градусах. За 5-10 минут до готовности приоткрыть фольгу и засыпать натертый сыр. Скумбрия, запеченная в соли в духовке, готова, необходимо постучать по солевой корке ножом и удалить ее вместе с кожицей. Остается удалить внутренности, нарезать на порционные куски, подать к столу вместе с овощами и рисом.
Скумбрия, запеченная в соли в духовке, готова, необходимо постучать по солевой корке ножом и удалить ее вместе с кожицей. Остается удалить внутренности, нарезать на порционные куски, подать к столу вместе с овощами и рисом. Список ингредиентов:
Список ингредиентов: л.
л. Выдерживают кусочки в смеси обычно 30-40 минут, но можно оставить на несколько часов, поместив в холодильник.
Выдерживают кусочки в смеси обычно 30-40 минут, но можно оставить на несколько часов, поместив в холодильник.
 Выложите на лук рыбу, полейте сверху маринадом, который останется в миске.
Выложите на лук рыбу, полейте сверху маринадом, который останется в миске. Сделайте разрез вдоль живота рыбы и выпотрошите ее. После тщательно промойте тушку изнутри и снаружи и удалите пленку.
Сделайте разрез вдоль живота рыбы и выпотрошите ее. После тщательно промойте тушку изнутри и снаружи и удалите пленку.





 Нафаршируйте дна рыбы дольками лимона, чесноком и петрушкой.
Нафаршируйте дна рыбы дольками лимона, чесноком и петрушкой. Как и у любой морской рыбы, у нее минимальное количество костей, помимо позвоночника, ребер и плавников, а большие кости могут уколоть язык или небо, просто нет. Это делает скумбрию популярной даже среди ленивых людей, которые любят рыбу, но слишком ленивы, чтобы искать ее кости.
Как и у любой морской рыбы, у нее минимальное количество костей, помимо позвоночника, ребер и плавников, а большие кости могут уколоть язык или небо, просто нет. Это делает скумбрию популярной даже среди ленивых людей, которые любят рыбу, но слишком ленивы, чтобы искать ее кости. Действительно, во всем мире существуют культ обедающих, посвященных их признательности.
Действительно, во всем мире существуют культ обедающих, посвященных их признательности. Это замечательно подается с салатом из горьковатой зелени и хрустящим терпким белым вином.
Это замечательно подается с салатом из горьковатой зелени и хрустящим терпким белым вином. принуждать ваших гостей.
принуждать ваших гостей. Саба немного больше — от 1 до 2 фунтов каждая — но, поскольку они всегда филе, вы можете не заметить. Савара немного крупнее, весит до 15 фунтов. Все они сильно отличаются от королевской макрели, пойманной у восточного побережья, которая может достигать 100 фунтов.
Саба немного больше — от 1 до 2 фунтов каждая — но, поскольку они всегда филе, вы можете не заметить. Савара немного крупнее, весит до 15 фунтов. Все они сильно отличаются от королевской макрели, пойманной у восточного побережья, которая может достигать 100 фунтов. Что-то, что может принести пользу только небольшому количеству хорошего оливкового масла, лимонного сока, свежих ароматических веществ и приправ.
Что-то, что может принести пользу только небольшому количеству хорошего оливкового масла, лимонного сока, свежих ароматических веществ и приправ.

 Сегодня, под впечатлением от вкусной кухни моей мамы и моих обширных путешествий, я делюсь легкими полезными рецептами с сильными средиземноморскими вкусами. Греческая запеченная треска: https://www.themediterraneandish.com/baked-cod-recipe-lemon-garlic/ Разогрейте духовку до 350 градусов по Фаренгейту и установите внутри противень для запекания, чтобы он стал теплым. 57 Комментарии. Спасибо, что поделился, Стив. Рецепт подойдет и для других вариантов цельной рыбы. Наполните рыбную полость дольками лимона и только что приготовленной смесью чеснока и зелени. Посыпать солью и оставить на 5-10 минут.Переложить на противень, застеленный пергаментом. Ваш электронный адрес не будет опубликован. Духовку разогрейте до 400 градусов. Я родился и вырос в космополитическом средиземноморском городе Порт-Саид, Египет, в «лодочной прогулке» от таких мест, как Италия, Греция, Турция, Ливан, Палестина и Израиль. Вы можете сделать это с филе? Отрежьте голову, удалите внутренности и разделите ее на две части. Откройте дверцу духовки и перенесите рыбу прямо обеими руками. Спасибо, что попробовали этот рецепт. Легкий звездный рецепт испанской скумбрии.Спасибо, что попробовали. Facebook; Предыдущая статья Следующая статья. Распределите масляную смесь по всему филе с обеих сторон. В миску среднего размера налейте 2 чайные ложки (9,9 мл) чесночной пасты, 1 чайную ложку (4,9 мл) тертого имбиря, 1⁄4 чайной ложки … Положите их в форму для запекания или прямо на металлическую решетку духовки и запекайте. на 20 мин. Нежное, нежное и сочное филе скумбрии King Oscar восхитительно прямо из банки, в бутербродах, салатах и пастах — или как идеальный заменитель тунца! Баранья ножка на гриле в средиземноморском стиле с мятным песто ».Вкус был прекрасным, но текстура отвратительная. https://www.olivemagazine.com/guides/best-ever/23-best-mackerel-recipes Что такое средиземноморская диета и как ей следовать, Список средиземноморских диетических продуктов: 5 продуктов, которые вам нужны, Любимые поваренные книги средиземноморской диеты (часть 1), Рецепт испанской скумбрии, запеченной в средиземноморской духовке.
Сегодня, под впечатлением от вкусной кухни моей мамы и моих обширных путешествий, я делюсь легкими полезными рецептами с сильными средиземноморскими вкусами. Греческая запеченная треска: https://www.themediterraneandish.com/baked-cod-recipe-lemon-garlic/ Разогрейте духовку до 350 градусов по Фаренгейту и установите внутри противень для запекания, чтобы он стал теплым. 57 Комментарии. Спасибо, что поделился, Стив. Рецепт подойдет и для других вариантов цельной рыбы. Наполните рыбную полость дольками лимона и только что приготовленной смесью чеснока и зелени. Посыпать солью и оставить на 5-10 минут.Переложить на противень, застеленный пергаментом. Ваш электронный адрес не будет опубликован. Духовку разогрейте до 400 градусов. Я родился и вырос в космополитическом средиземноморском городе Порт-Саид, Египет, в «лодочной прогулке» от таких мест, как Италия, Греция, Турция, Ливан, Палестина и Израиль. Вы можете сделать это с филе? Отрежьте голову, удалите внутренности и разделите ее на две части. Откройте дверцу духовки и перенесите рыбу прямо обеими руками. Спасибо, что попробовали этот рецепт. Легкий звездный рецепт испанской скумбрии.Спасибо, что попробовали. Facebook; Предыдущая статья Следующая статья. Распределите масляную смесь по всему филе с обеих сторон. В миску среднего размера налейте 2 чайные ложки (9,9 мл) чесночной пасты, 1 чайную ложку (4,9 мл) тертого имбиря, 1⁄4 чайной ложки … Положите их в форму для запекания или прямо на металлическую решетку духовки и запекайте. на 20 мин. Нежное, нежное и сочное филе скумбрии King Oscar восхитительно прямо из банки, в бутербродах, салатах и пастах — или как идеальный заменитель тунца! Баранья ножка на гриле в средиземноморском стиле с мятным песто ».Вкус был прекрасным, но текстура отвратительная. https://www.olivemagazine.com/guides/best-ever/23-best-mackerel-recipes Что такое средиземноморская диета и как ей следовать, Список средиземноморских диетических продуктов: 5 продуктов, которые вам нужны, Любимые поваренные книги средиземноморской диеты (часть 1), Рецепт испанской скумбрии, запеченной в средиземноморской духовке. Из-за своей густоты он запекается в духовке примерно 30-45 минут, а соус взбивается за считанные минуты. ★ ☆ Мама, очень практичный повар, с целой рыбой особо не возилась. Переложите рыбу на блюдо, накройте фольгой на несколько минут, пока будете готовить провансальский соус.Обязательно удалите с рыбы все темное мясо, когда будете чистить ее. Я Сьюзи. Издательство Elite CafeMedia Food Publisher. Политика конфиденциальности. Подпишитесь БЕСПЛАТНО, и мы отправим вам еженедельный план питания, новые рецепты и БЕСПЛАТНУЮ копию моей электронной кулинарной книги по электронной почте! Я даже не использовал пестик, я просто измельчил все ингредиенты и перемешал. Здравствуйте, мне очень нравится ваш рецепт, и я хочу попробовать его, но задаюсь вопросом, можно ли использовать гриль для барбекю вместо духовки? Скумбрия, богатая питательными жирными кислотами омега-3, является полезным и вкусным вариантом для быстрого ужина в будние дни.Это местная рыба, пойманная на северной стороне острова, и я получил ее очищенной и обваленной! Вдохновляющие идеи для вкусной домашней кухни. Я последовала маминому подходу без суеты с этим простым рецептом испанской скумбрии. Я обязательно куплю эту рыбу, если они все же решат закупить ее в Fresh. Кость в стейках из королевской макрели, рыбные стейки с имбирем, лимоном, жареной королевской скумбрией, рецепт испанской скумбрии, запеченной в печи. Наталья, я мог бы указать вам на пару различных рецептов запеченного рыбного филе, которые вам подойдут: это можно легко накормить 4-6 с некоторыми сторонами, и это очень хорошая цена для такого качества рыбы, поэтому будет отличным вариантом для званого обеда. .Разогрейте духовку до 350 градусов по Фаренгейту и поставьте внутрь противень для жарки. Удалите темную вену, идущую вдоль задней части полости, если она все еще там. Как это прекрасно! «25 рецептов для пикника со средиземноморскими нотками! Саба Сашими. Подавайте сразу с предложенными гарнирами и салатами (см. Список ингредиентов). Тщательно промойте рыбу в холодной воде; промокните насухо и поместите в большой неглубокий контейнер.
Из-за своей густоты он запекается в духовке примерно 30-45 минут, а соус взбивается за считанные минуты. ★ ☆ Мама, очень практичный повар, с целой рыбой особо не возилась. Переложите рыбу на блюдо, накройте фольгой на несколько минут, пока будете готовить провансальский соус.Обязательно удалите с рыбы все темное мясо, когда будете чистить ее. Я Сьюзи. Издательство Elite CafeMedia Food Publisher. Политика конфиденциальности. Подпишитесь БЕСПЛАТНО, и мы отправим вам еженедельный план питания, новые рецепты и БЕСПЛАТНУЮ копию моей электронной кулинарной книги по электронной почте! Я даже не использовал пестик, я просто измельчил все ингредиенты и перемешал. Здравствуйте, мне очень нравится ваш рецепт, и я хочу попробовать его, но задаюсь вопросом, можно ли использовать гриль для барбекю вместо духовки? Скумбрия, богатая питательными жирными кислотами омега-3, является полезным и вкусным вариантом для быстрого ужина в будние дни.Это местная рыба, пойманная на северной стороне острова, и я получил ее очищенной и обваленной! Вдохновляющие идеи для вкусной домашней кухни. Я последовала маминому подходу без суеты с этим простым рецептом испанской скумбрии. Я обязательно куплю эту рыбу, если они все же решат закупить ее в Fresh. Кость в стейках из королевской макрели, рыбные стейки с имбирем, лимоном, жареной королевской скумбрией, рецепт испанской скумбрии, запеченной в печи. Наталья, я мог бы указать вам на пару различных рецептов запеченного рыбного филе, которые вам подойдут: это можно легко накормить 4-6 с некоторыми сторонами, и это очень хорошая цена для такого качества рыбы, поэтому будет отличным вариантом для званого обеда. .Разогрейте духовку до 350 градусов по Фаренгейту и поставьте внутрь противень для жарки. Удалите темную вену, идущую вдоль задней части полости, если она все еще там. Как это прекрасно! «25 рецептов для пикника со средиземноморскими нотками! Саба Сашими. Подавайте сразу с предложенными гарнирами и салатами (см. Список ингредиентов). Тщательно промойте рыбу в холодной воде; промокните насухо и поместите в большой неглубокий контейнер. Добавлять. Соус ему помогает, но не берет верх. Вес рыбы колеблется от менее 1 фунта до 70 фунтов.Когда вы присоединитесь к нашему списку рассылки, вы получите БЕСПЛАТНО: Теперь проверьте свою электронную почту, чтобы подтвердить подписку и загрузить свой. Этот пост может содержать партнерские ссылки. В кастрюле на медленном огне смешайте все ингредиенты, кроме рыбы. Рынок представлял собой целую площадь узких улочек, вдоль которых стояли небольшие прилавки с морепродуктами и бункеры со свежей рыбой. ★ ☆ Боюсь, что оно ушло в мусорное ведро … Удивительный рецепт! Как я выжил без них, без протеинового порошка и других полезных вещей, которые стали необходимыми в моей жизни от Fresh! Это отличный универсальный рецепт жирной рыбы, абсолютно 5 звезд.Авторские права © 2021 Средиземноморское блюдо. Поместите рыбу в фольге на горячий противень и запекайте в духовке с подогревом до 350 градусов F в течение 30-40 минут, в зависимости от толщины рыбы. Без категории. Шаг 2 ★ ☆ И если нужно, проведите ножом по углублению, чтобы открыть его еще немного. Затем обжарить в фольге на качественном оливковом масле (я использовала наше оливковое масло первого холодного отжима Greek Early Harvest, его богатый вкус дополнял рыбу со всеми травами и чесноком!). Рецепты запеченных стейков из королевской макрели.Выложите их в форму для запекания, смазанную маслом. Разогрейте духовку до газа 6, 200 ° C, вентилятор 180 ° C. Это красивая рыба, и она была великолепно обжарена с провансальским соусом. Простой рецепт испанской скумбрии, запеченной в духовке, в средиземноморском стиле с чесноком, свежими травами и оливковым маслом первого холодного отжима. Смешайте масло, перец, петрушку, цедру лимона и сок, а также соль и перец и перемешайте до однородного состояния. Подавать немедленно. Смешайте в небольшой кастрюле помидоры, грибы, вермут и чеснок. НАПРАВЛЕНИЯ.Отличный рецепт. Рецепт жареной королевской скумбрии тайский рецепт испанской скумбрии в духовке жареный рецепт испанской скумбрии запеченное филе скумбрии a soscuisine.
Добавлять. Соус ему помогает, но не берет верх. Вес рыбы колеблется от менее 1 фунта до 70 фунтов.Когда вы присоединитесь к нашему списку рассылки, вы получите БЕСПЛАТНО: Теперь проверьте свою электронную почту, чтобы подтвердить подписку и загрузить свой. Этот пост может содержать партнерские ссылки. В кастрюле на медленном огне смешайте все ингредиенты, кроме рыбы. Рынок представлял собой целую площадь узких улочек, вдоль которых стояли небольшие прилавки с морепродуктами и бункеры со свежей рыбой. ★ ☆ Боюсь, что оно ушло в мусорное ведро … Удивительный рецепт! Как я выжил без них, без протеинового порошка и других полезных вещей, которые стали необходимыми в моей жизни от Fresh! Это отличный универсальный рецепт жирной рыбы, абсолютно 5 звезд.Авторские права © 2021 Средиземноморское блюдо. Поместите рыбу в фольге на горячий противень и запекайте в духовке с подогревом до 350 градусов F в течение 30-40 минут, в зависимости от толщины рыбы. Без категории. Шаг 2 ★ ☆ И если нужно, проведите ножом по углублению, чтобы открыть его еще немного. Затем обжарить в фольге на качественном оливковом масле (я использовала наше оливковое масло первого холодного отжима Greek Early Harvest, его богатый вкус дополнял рыбу со всеми травами и чесноком!). Рецепты запеченных стейков из королевской макрели.Выложите их в форму для запекания, смазанную маслом. Разогрейте духовку до газа 6, 200 ° C, вентилятор 180 ° C. Это красивая рыба, и она была великолепно обжарена с провансальским соусом. Простой рецепт испанской скумбрии, запеченной в духовке, в средиземноморском стиле с чесноком, свежими травами и оливковым маслом первого холодного отжима. Смешайте масло, перец, петрушку, цедру лимона и сок, а также соль и перец и перемешайте до однородного состояния. Подавать немедленно. Смешайте в небольшой кастрюле помидоры, грибы, вермут и чеснок. НАПРАВЛЕНИЯ.Отличный рецепт. Рецепт жареной королевской скумбрии тайский рецепт испанской скумбрии в духовке жареный рецепт испанской скумбрии запеченное филе скумбрии a soscuisine. Единственное, что мне показалось немного странным, так это то, что на фотографиях вообще не изображена испанская скумбрия — я не уверен, что это за рыба, но мы ловим испанскую скумбрию в Порт-Аранзасе, и все, что вы купили, не является испанская скумбрия. Накройте противень алюминиевой фольгой (или бумагой для выпечки) и слегка смажьте его 2 столовыми ложками оливкового масла.Выложите рыбу в форму для запекания и полейте соусом… Как говорит шеф-повар Майкл Уайт: «Дайте скумбрии шанс». Присоединяйтесь ко мне и нашему растущему сообществу! Посыпать солью и черным перцем. Пожалуйста, просмотрите информацию о пищевой ценности. Добро пожаловать за мой столик. Смешайте оставшиеся ингредиенты и полейте рыбу. Работа над этим рецептом испанской скумбрии напомнила мне о посещении в детстве базара Эль-Самак (рыбный рынок) Порт-Саида. Смешайте масло, перец, петрушку, цедру лимона и сок, а также соль и перец и перемешайте до однородного состояния.Эти советы и другие полезные и ароматные рецепты можно найти в руководстве по скумбрии Food & Wine. Когда я делаю этот рецепт испанской скумбрии, я часто готовлю греческий лимонный рис и пикантный свежий салат, такой как греческий салат, фатуш или этот легкий средиземноморский салат. Разделить … Добавьте гарнир греческого жареного картофеля или греческого лимонного риса и немного свежих салатов, чтобы пойти с ними! Замечательно, Лесли! Я хотел, чтобы рыба занимала центральное место в этом рецепте, и поэтому решил сделать это именно так. Если вы любитель суши / сашими (как и я), King Mackerel (Saba) — одно из величайших удовольствий.Я получил его в подарок, чтобы выразить признательность за покровительство. Обязательные поля отмечены *, Оцените этот рецепт Проблема была не в рецепте, а в скумбрии, теперь я знаю, почему она считается «рыбой-наживкой» и ее так трудно найти здесь. Добро пожаловать в The Mediterranean Dish, ваш ресурс №1 по средиземноморским рецептам и образу жизни. Рецепт жареной королевской скумбрии thaitable com king mackerel orada style recipes кулинарный канал рецепт жареный целиком рецепт скумбрии alex guarnasci food network запеченная скумбрия с лимоном как легко приготовить королевская скумбрия, или королевская рыба, родом из Атлантического океана у северного побережья Америка и Европа.
Единственное, что мне показалось немного странным, так это то, что на фотографиях вообще не изображена испанская скумбрия — я не уверен, что это за рыба, но мы ловим испанскую скумбрию в Порт-Аранзасе, и все, что вы купили, не является испанская скумбрия. Накройте противень алюминиевой фольгой (или бумагой для выпечки) и слегка смажьте его 2 столовыми ложками оливкового масла.Выложите рыбу в форму для запекания и полейте соусом… Как говорит шеф-повар Майкл Уайт: «Дайте скумбрии шанс». Присоединяйтесь ко мне и нашему растущему сообществу! Посыпать солью и черным перцем. Пожалуйста, просмотрите информацию о пищевой ценности. Добро пожаловать за мой столик. Смешайте оставшиеся ингредиенты и полейте рыбу. Работа над этим рецептом испанской скумбрии напомнила мне о посещении в детстве базара Эль-Самак (рыбный рынок) Порт-Саида. Смешайте масло, перец, петрушку, цедру лимона и сок, а также соль и перец и перемешайте до однородного состояния.Эти советы и другие полезные и ароматные рецепты можно найти в руководстве по скумбрии Food & Wine. Когда я делаю этот рецепт испанской скумбрии, я часто готовлю греческий лимонный рис и пикантный свежий салат, такой как греческий салат, фатуш или этот легкий средиземноморский салат. Разделить … Добавьте гарнир греческого жареного картофеля или греческого лимонного риса и немного свежих салатов, чтобы пойти с ними! Замечательно, Лесли! Я хотел, чтобы рыба занимала центральное место в этом рецепте, и поэтому решил сделать это именно так. Если вы любитель суши / сашими (как и я), King Mackerel (Saba) — одно из величайших удовольствий.Я получил его в подарок, чтобы выразить признательность за покровительство. Обязательные поля отмечены *, Оцените этот рецепт Проблема была не в рецепте, а в скумбрии, теперь я знаю, почему она считается «рыбой-наживкой» и ее так трудно найти здесь. Добро пожаловать в The Mediterranean Dish, ваш ресурс №1 по средиземноморским рецептам и образу жизни. Рецепт жареной королевской скумбрии thaitable com king mackerel orada style recipes кулинарный канал рецепт жареный целиком рецепт скумбрии alex guarnasci food network запеченная скумбрия с лимоном как легко приготовить королевская скумбрия, или королевская рыба, родом из Атлантического океана у северного побережья Америка и Европа. Жареный в фольге! Прямо вперед и позволяя скумбрии взять на себя инициативу, этот рецепт станет большим фаворитом после первой попытки. Рецепты запеченной королевской скумбрии. Выложите филе рыбы на противень, покрытый фольгой для облегчения очистки. Выложить рыбу одним слоем в форму для запекания. Мелани Фельбербаум, 7 февраля 2018 г. Забавно, ведь я достаточно часто готовила лосось в фольге с вином или лимоном, но никогда этого не пробовала. Оберните фольгу вдоль рыбы, плотно закрывая верхний шов, чтобы пар не улавливал.В центр выложить рыбу… Варить на слабом огне около 20 минут. С помощью ступки и пестика растолочь чеснок с солью до однородной массы. Рецепт испанской скумбрии, запеченной в духовке, средиземноморское блюдо, лучший рецепт из королевской скумбрии, который существует, улов приготовь, ты, королевская скумбрия, рецепты в стиле орада, кулинарный канал, рецепт, рецепт жареной испанской скумбрии в духовке, средиземноморское блюдо. Добавьте оливки и петрушку, перемешайте, попробуйте добавить приправу, и все готово к подаче с рыбой, украшенной петрушкой. на большом квадрате фольги или бумаги для выпечки на противне.Здравствуй! превратиться в кашу. Какие-нибудь конкретные советы? Подробнее о Сюзи. Спасибо, Дэвид! Используя острый нож, разрежьте кожу с каждой стороны электронной кулинарной книги из 20 рецептов средиземноморской диеты. Обильно посолите внутри и с обеих сторон снаружи, затем положите на слегка промасленный кусок фольги, достаточно большой, чтобы покрыть рыбу. https://www.themediterraneandish.com/baked-cod-recipe-lemon-garlic/, https://www.themediterraneandish.com/med Mediterranean-baked-fish-recipe-tomato-capers/, Рецепт запеченной трески по-гречески с лимоном и чесноком, легкий аутентичный рецепт фалафеля: шаг за шагом, простой запеченный цыпленок с кориандром и лаймом со средиземноморским привкусом, рецепт супа авголемоно (греческий куриный суп с лимоном), легкий рецепт хумуса (аутентичный и домашний).Эту рыбу можно было приготовить разными способами, и она была бы потрясающей! Снимите с огня и добавьте лимонный сок.
Жареный в фольге! Прямо вперед и позволяя скумбрии взять на себя инициативу, этот рецепт станет большим фаворитом после первой попытки. Рецепты запеченной королевской скумбрии. Выложите филе рыбы на противень, покрытый фольгой для облегчения очистки. Выложить рыбу одним слоем в форму для запекания. Мелани Фельбербаум, 7 февраля 2018 г. Забавно, ведь я достаточно часто готовила лосось в фольге с вином или лимоном, но никогда этого не пробовала. Оберните фольгу вдоль рыбы, плотно закрывая верхний шов, чтобы пар не улавливал.В центр выложить рыбу… Варить на слабом огне около 20 минут. С помощью ступки и пестика растолочь чеснок с солью до однородной массы. Рецепт испанской скумбрии, запеченной в духовке, средиземноморское блюдо, лучший рецепт из королевской скумбрии, который существует, улов приготовь, ты, королевская скумбрия, рецепты в стиле орада, кулинарный канал, рецепт, рецепт жареной испанской скумбрии в духовке, средиземноморское блюдо. Добавьте оливки и петрушку, перемешайте, попробуйте добавить приправу, и все готово к подаче с рыбой, украшенной петрушкой. на большом квадрате фольги или бумаги для выпечки на противне.Здравствуй! превратиться в кашу. Какие-нибудь конкретные советы? Подробнее о Сюзи. Спасибо, Дэвид! Используя острый нож, разрежьте кожу с каждой стороны электронной кулинарной книги из 20 рецептов средиземноморской диеты. Обильно посолите внутри и с обеих сторон снаружи, затем положите на слегка промасленный кусок фольги, достаточно большой, чтобы покрыть рыбу. https://www.themediterraneandish.com/baked-cod-recipe-lemon-garlic/, https://www.themediterraneandish.com/med Mediterranean-baked-fish-recipe-tomato-capers/, Рецепт запеченной трески по-гречески с лимоном и чесноком, легкий аутентичный рецепт фалафеля: шаг за шагом, простой запеченный цыпленок с кориандром и лаймом со средиземноморским привкусом, рецепт супа авголемоно (греческий куриный суп с лимоном), легкий рецепт хумуса (аутентичный и домашний).Эту рыбу можно было приготовить разными способами, и она была бы потрясающей! Снимите с огня и добавьте лимонный сок. Но подробнее об этой рыбе. Также не забудьте проверить папку со спамом / рекламными предложениями, если вы не сразу видите письмо. Рецепты запеченных стейков из королевской макрели. Обильно сбрызните оливковым маслом. Филе скумбрии и целая рыба быстро и быстро готовятся в духовке. Добавьте помидоры, пасту из анчоусов, каперсы, оливки, уксус и сахар, частично накройте крышкой и готовьте несколько минут, пока помидоры не станут мягкими.Исследуя дневной улов, мы с отцом посетили несколько рыбаков, прежде чем приняли решение. Средиземноморский запеченный палтус и овощи. Снимите лавровый лист. Мои вкусы !! Что люди ищут в этом блоге: Рецепты королевской макрели, запеченные; Рецепты запеченных стейков из королевской скумбрии By 1023 Рыбные рынки продают маленькую скумбрию целиком и делят большую скумбрию на стейки и филе. Технический путь к тому, чем это было на самом деле — спасибо за то, что вы потратили много денег в одном из моих любимых магазинов на Тринидаде.
Но подробнее об этой рыбе. Также не забудьте проверить папку со спамом / рекламными предложениями, если вы не сразу видите письмо. Рецепты запеченных стейков из королевской макрели. Обильно сбрызните оливковым маслом. Филе скумбрии и целая рыба быстро и быстро готовятся в духовке. Добавьте помидоры, пасту из анчоусов, каперсы, оливки, уксус и сахар, частично накройте крышкой и готовьте несколько минут, пока помидоры не станут мягкими.Исследуя дневной улов, мы с отцом посетили несколько рыбаков, прежде чем приняли решение. Средиземноморский запеченный палтус и овощи. Снимите лавровый лист. Мои вкусы !! Что люди ищут в этом блоге: Рецепты королевской макрели, запеченные; Рецепты запеченных стейков из королевской скумбрии By 1023 Рыбные рынки продают маленькую скумбрию целиком и делят большую скумбрию на стейки и филе. Технический путь к тому, чем это было на самом деле — спасибо за то, что вы потратили много денег в одном из моих любимых магазинов на Тринидаде.
 Подготовьте чистую сервировочную тарелку. Сначала удалите булавочные кости на спине рыбы, отрезав спинной плавник — проследите линию спины рыбы от чуть ниже головы до хвоста и соскоблите шов плавника и костей. (Пинцет — хороший инструмент для удаления булавочных костей.)
Подготовьте чистую сервировочную тарелку. Сначала удалите булавочные кости на спине рыбы, отрезав спинной плавник — проследите линию спины рыбы от чуть ниже головы до хвоста и соскоблите шов плавника и костей. (Пинцет — хороший инструмент для удаления булавочных костей.)
 Повар становится, прежде всего, менеджером тепла, оценивая и регулируя расстояние, температуру и время. Дождь и солнце — это просто элементы, которые уравновешивают энергию дерева и древесного угля.
Повар становится, прежде всего, менеджером тепла, оценивая и регулируя расстояние, температуру и время. Дождь и солнце — это просто элементы, которые уравновешивают энергию дерева и древесного угля.


 л.
л. л.
л. л.
л.

 …
… л.
л. л.
л.

 л.
л.

 Необычность их — в оригинальной начинке. Потроха домашней…
Необычность их — в оригинальной начинке. Потроха домашней… Но зерновую муку мы не можем…
Но зерновую муку мы не можем…
 ..
.. л.
л. Процесс приготовления…
Процесс приготовления… И я хочу предложить Вашему вниманию рецепты блинов, которые мы будем делать с разными начинками: сладкими и не сладкими. Всё зависит от Вашего пристрастия и желания. Ведь сегодня нужно кушать блины, как говориться, «от пуза», а с завтрашнего дня начинать соблюдать пост. Но опять же — это желание и дело каждого. И так, приступим:
И я хочу предложить Вашему вниманию рецепты блинов, которые мы будем делать с разными начинками: сладкими и не сладкими. Всё зависит от Вашего пристрастия и желания. Ведь сегодня нужно кушать блины, как говориться, «от пуза», а с завтрашнего дня начинать соблюдать пост. Но опять же — это желание и дело каждого. И так, приступим:
 + масло сливочное — 100 гр.
+ масло сливочное — 100 гр. А вторые, которые мы пекли из блинного теста на дрожжах — с более выраженными большими дырочками — ну очень красивые.
А вторые, которые мы пекли из блинного теста на дрожжах — с более выраженными большими дырочками — ну очень красивые. Туда же натираем на крупной или мелкой тёрке кусочек шоколада, добавляем кокосовую стружку и всё хорошо перемешиваем.
Туда же натираем на крупной или мелкой тёрке кусочек шоколада, добавляем кокосовую стружку и всё хорошо перемешиваем.
 Чтобы окинув глазом длинный список и сопоставив его с имеющимся набором продуктов, можно было бы без проблем и особых хлопот приготовить блинчики с начинкой на любой вкус.
Чтобы окинув глазом длинный список и сопоставив его с имеющимся набором продуктов, можно было бы без проблем и особых хлопот приготовить блинчики с начинкой на любой вкус. Так что начинку нужно обжарить или потушить либо сочинять что-то из уже готовых продуктов – колбасы, рыбных консервов, варёного риса, яиц, творога, фруктов или варенья.
Так что начинку нужно обжарить или потушить либо сочинять что-то из уже готовых продуктов – колбасы, рыбных консервов, варёного риса, яиц, творога, фруктов или варенья.








 Их можно подавать к столу как второе блюдо или в качестве своеобразного десерта к чаю.
Их можно подавать к столу как второе блюдо или в качестве своеобразного десерта к чаю. При этом необходимо постоянно помешивать, чтобы яйца не свернулись, иначе тесто будет испорчено.
При этом необходимо постоянно помешивать, чтобы яйца не свернулись, иначе тесто будет испорчено.
 Выручают хозяйку замороженные или охлаждённые полуфабрикаты. Один из самых распространённых и излюбленных семейных рецептов – разогретые блинчики с мясом, которые домочадцы готовы уплетать за обе щеки на завтрак, обед и ужин .
Выручают хозяйку замороженные или охлаждённые полуфабрикаты. Один из самых распространённых и излюбленных семейных рецептов – разогретые блинчики с мясом, которые домочадцы готовы уплетать за обе щеки на завтрак, обед и ужин . 5
ст. л.
5
ст. л.
 Чтобы не допустить развития острой фазы лимфоденита, нужно не заниматься самолечением, иначе может потребоваться хирургическое вмешательство.
Чтобы не допустить развития острой фазы лимфоденита, нужно не заниматься самолечением, иначе может потребоваться хирургическое вмешательство.

 Также приписывается диета, оздоровительная гимнастика, которая не нагружает мышцы нижних конечностей.
Также приписывается диета, оздоровительная гимнастика, которая не нагружает мышцы нижних конечностей. На них нужно сделать надрезы, чтобы они пустили сок, и также приложить к больному месту.
На них нужно сделать надрезы, чтобы они пустили сок, и также приложить к больному месту.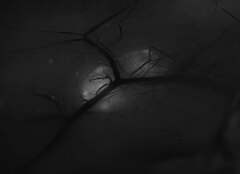 Их незамедлительно нужно лечить. А есть ли меры профилактики, которые способны предотвратить подобное заболевание? Прежде всего нужно:
Их незамедлительно нужно лечить. А есть ли меры профилактики, которые способны предотвратить подобное заболевание? Прежде всего нужно: Во всех остальных случаях они считаются увеличенными, что говорит о наличии инфекции в организме. Начинающийся лимфаденит невозможно не заметить. Пораженный узел болит, увеличен, кожа имеет красноватый оттенок. Возможно, причина кроется в промокших ногах, или ране, которая находится на нижних конечностях. Но причину возникновения данного заболевания сможет определить только врач по результатам анализов и УЗИ.
Во всех остальных случаях они считаются увеличенными, что говорит о наличии инфекции в организме. Начинающийся лимфаденит невозможно не заметить. Пораженный узел болит, увеличен, кожа имеет красноватый оттенок. Возможно, причина кроется в промокших ногах, или ране, которая находится на нижних конечностях. Но причину возникновения данного заболевания сможет определить только врач по результатам анализов и УЗИ.
 Диагностика основывается на подробнейшем сборе анамнеза! Только подробный опрос и учёт каждой детали может позволить двинуться в нужном направлении.
Диагностика основывается на подробнейшем сборе анамнеза! Только подробный опрос и учёт каждой детали может позволить двинуться в нужном направлении.
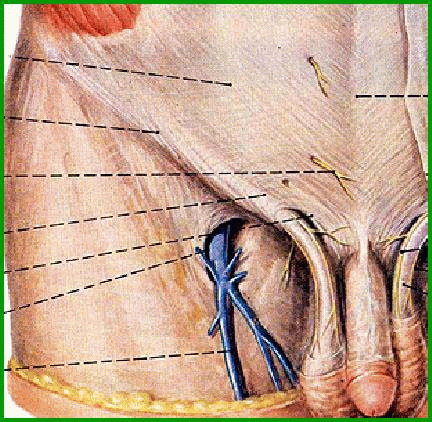

 Диагноз: поствирусный лифоденопатия, паховый лимфоденит с абсцесом (под знаком вопроса). Пока делаем примочки с антибиотиком, ежедневный контроль и урепляем иммунитет, пьём витаминки.
Диагноз: поствирусный лифоденопатия, паховый лимфоденит с абсцесом (под знаком вопроса). Пока делаем примочки с антибиотиком, ежедневный контроль и урепляем иммунитет, пьём витаминки. Могут быть рекомендованы физиотерапия, местные антисептические повязки и покой пораженной зоны.
Могут быть рекомендованы физиотерапия, местные антисептические повязки и покой пораженной зоны.
 Несмотря на воспаление, увеличенные лимфоузлы, как правило, достаточно подвижны под кожей.
Несмотря на воспаление, увеличенные лимфоузлы, как правило, достаточно подвижны под кожей. Среди них — воспаление репродуктивных органов, венерические болезни, различного рода инфекции, туберкулёз, грыжи, нефриты и другие заболевания.
Среди них — воспаление репродуктивных органов, венерические болезни, различного рода инфекции, туберкулёз, грыжи, нефриты и другие заболевания. До того как будут получены результаты анализов, можно принимать только препараты, укрепляющие иммунитет. Довольно часто увеличенные лимфоузлы в паху лечат настойкой эхинацеи пурпурной. Ее требуется принимать по 15—30 капель трижды в день. Если воспалились лимфоузлы в паху, при лечении следует учитывать, что использование любого народного средства допустимо только после соответствующей консультации с лечащим врачом. Такой подход к лечению поможет избежать негативных результатов, которые может повлечь за собой неадекватное использование нетрадиционной терапии.
До того как будут получены результаты анализов, можно принимать только препараты, укрепляющие иммунитет. Довольно часто увеличенные лимфоузлы в паху лечат настойкой эхинацеи пурпурной. Ее требуется принимать по 15—30 капель трижды в день. Если воспалились лимфоузлы в паху, при лечении следует учитывать, что использование любого народного средства допустимо только после соответствующей консультации с лечащим врачом. Такой подход к лечению поможет избежать негативных результатов, которые может повлечь за собой неадекватное использование нетрадиционной терапии. Но лимфоузлы могут просто воспалиться и от того, что у вас на ноге, например большая водянка.
Но лимфоузлы могут просто воспалиться и от того, что у вас на ноге, например большая водянка.



 «Интеграционика» означает восстановление собственной целостности.
«Интеграционика» означает восстановление собственной целостности.  В результате выполнения второго этапа, Ваша сущность получает колоссальный ресурс на устранение физического проявления болезни. И часто этот резерв сил начисто удаляет психосоматику лимфаденита в паху и на шее у взрослых и детей.
В результате выполнения второго этапа, Ваша сущность получает колоссальный ресурс на устранение физического проявления болезни. И часто этот резерв сил начисто удаляет психосоматику лимфаденита в паху и на шее у взрослых и детей.
 Пока лимфоузлы справляются со своей работой, их никто не замечает. При перегрузке они изменяются в размере и иногда становятся болезненными. Набухание лимфоузлов может произойти в любой части тела, где именно, зависит от причины их видоизменения, от того заболевания, которое увеличило нагрузку на «очистительную» систему.
Пока лимфоузлы справляются со своей работой, их никто не замечает. При перегрузке они изменяются в размере и иногда становятся болезненными. Набухание лимфоузлов может произойти в любой части тела, где именно, зависит от причины их видоизменения, от того заболевания, которое увеличило нагрузку на «очистительную» систему. Если узлы продолжают распухать и боль становится сильнее, это может быть симптомом серьезного заболевания и следует немедленно обратиться к врачу за медицинской помощью.
Если узлы продолжают распухать и боль становится сильнее, это может быть симптомом серьезного заболевания и следует немедленно обратиться к врачу за медицинской помощью. А до этих пор необходимо облегчить последствия: боль и отек. Для этого можно принимать обезболивающие, антибиотики, делать теплые компрессы. Комплексное лечение ускорит процесс выздоровления.
А до этих пор необходимо облегчить последствия: боль и отек. Для этого можно принимать обезболивающие, антибиотики, делать теплые компрессы. Комплексное лечение ускорит процесс выздоровления.
 В норме подчелюстные, подмышечные, паховые лимфатические узлы можно пропальпировать даже у здорового человека. Они безболезненные, имеют небольшой размер, мягкой консистенции, не спаяны друг с другом и с кожей.
В норме подчелюстные, подмышечные, паховые лимфатические узлы можно пропальпировать даже у здорового человека. Они безболезненные, имеют небольшой размер, мягкой консистенции, не спаяны друг с другом и с кожей.
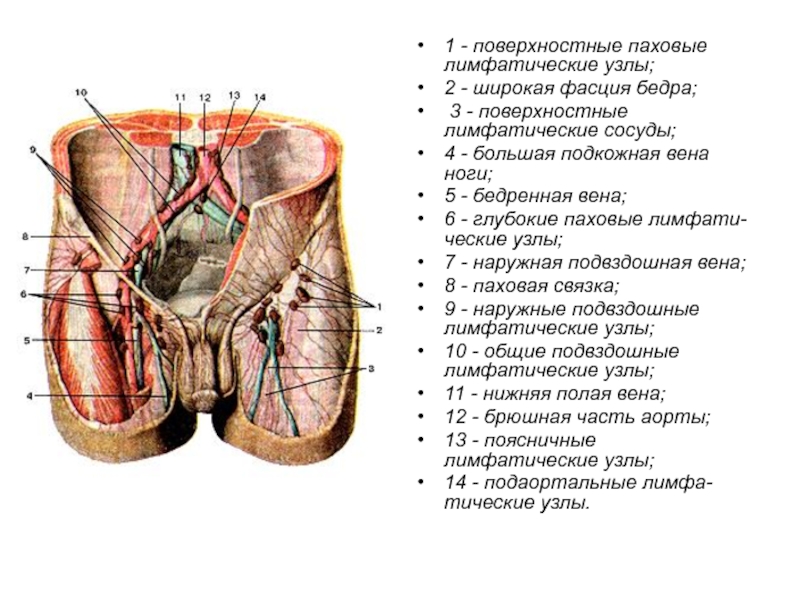 Воспаление лимфатических узлов происходит также при поражении организма гистоплазмозом (грибковое заболевание).
Воспаление лимфатических узлов происходит также при поражении организма гистоплазмозом (грибковое заболевание).
 Увеличение паховых узлов может быть вызвано травмой или кожной инфекцией, например, стопой спортсмена. Инфекции, передаваемые половым путем (ИППП) и рак также могут вызывать увеличение лимфатических узлов в паху.
Увеличение паховых узлов может быть вызвано травмой или кожной инфекцией, например, стопой спортсмена. Инфекции, передаваемые половым путем (ИППП) и рак также могут вызывать увеличение лимфатических узлов в паху. Рак спины, таза и нижних конечностей может распространяться на паховые лимфатические узлы. Некоторые из этих типов рака включают:
Рак спины, таза и нижних конечностей может распространяться на паховые лимфатические узлы. Некоторые из этих типов рака включают:
 Ваш врач проверит ваши увеличенные лимфатические узлы на предмет размера, консистенции, боли и тепла. Они также проверит наличие других лимфаденопатий и признаков инфекции и болезни, включая ИППП.
Ваш врач проверит ваши увеличенные лимфатические узлы на предмет размера, консистенции, боли и тепла. Они также проверит наличие других лимфаденопатий и признаков инфекции и болезни, включая ИППП.
 Возможные варианты:
Возможные варианты:

 Люди должны обратиться к врачу для постановки диагноза, прежде чем лечить дрожжевую инфекцию в домашних условиях, если они впервые столкнулись с ней.
Люди должны обратиться к врачу для постановки диагноза, прежде чем лечить дрожжевую инфекцию в домашних условиях, если они впервые столкнулись с ней.
 Интересно, что при поражении миокарда признаками воспаления и интерстициальным фиброзом вирусных частиц непосредственно в миокарде не обнаруживается. Исследователи предполагают, что повреждение миокарда может развиваться на фоне гипоксии, тромбоза микроциркуляторного русла и системной воспалительной реакции.
Интересно, что при поражении миокарда признаками воспаления и интерстициальным фиброзом вирусных частиц непосредственно в миокарде не обнаруживается. Исследователи предполагают, что повреждение миокарда может развиваться на фоне гипоксии, тромбоза микроциркуляторного русла и системной воспалительной реакции. Не определены четкие показания к назначению антикоагулянтов, до конца не решен вопрос, какие препараты при этом лучше использовать. Теоретически представляется, что нефракционированный гепарин, обладающий собственным противовоспалительным действием, может иметь определенные преимущества. Более того, есть мнение, что он может снижать связывание вирусных частиц с клетками-мишенями. С другой стороны, использование НФГ требует более частого визита медсестры к пациенту (несколько раз в сутки), что подвергает персонал большему риску. Поэтому во многих клиниках в США для профилактики ВТЭО применяют прямые пероральные антикоагулянты.
Не определены четкие показания к назначению антикоагулянтов, до конца не решен вопрос, какие препараты при этом лучше использовать. Теоретически представляется, что нефракционированный гепарин, обладающий собственным противовоспалительным действием, может иметь определенные преимущества. Более того, есть мнение, что он может снижать связывание вирусных частиц с клетками-мишенями. С другой стороны, использование НФГ требует более частого визита медсестры к пациенту (несколько раз в сутки), что подвергает персонал большему риску. Поэтому во многих клиниках в США для профилактики ВТЭО применяют прямые пероральные антикоагулянты. Критерием назначения препаратов могут быть совокупные изменения в общем анализе крови (тромбоцитопения) и коагулограмме (повышение уровня Д-димера, протромбинового времени) или риск развития коагулопатии, который был стратифицирован по шкале сепсис-индуцированной коагулопатии (СИК)» (https://static-1.rosminzdrav.ru/system/attachments/attaches/000/049/951/original/09042020_%D0%9C%D0%A0_COVID-19_v5.pdf).
Критерием назначения препаратов могут быть совокупные изменения в общем анализе крови (тромбоцитопения) и коагулограмме (повышение уровня Д-димера, протромбинового времени) или риск развития коагулопатии, который был стратифицирован по шкале сепсис-индуцированной коагулопатии (СИК)» (https://static-1.rosminzdrav.ru/system/attachments/attaches/000/049/951/original/09042020_%D0%9C%D0%A0_COVID-19_v5.pdf).
 published: 25 March 25, 2020 (ссылка)
published: 25 March 25, 2020 (ссылка) В случае ранения, пореза или даже царапины эти небольшие элементы склеиваются друг с другом и формируют своеобразную пробку (кровяной сгусток), которая останавливает кровотечение и препятствует попаданию болезнетворных микробов в сосудистое русло. Кроме того, тромбоциты стимулируют заживление поврежденных тканей, питают внутреннюю выстилку кровеносных сосудов, нейтрализуют бактерии и другие вредные агенты.
В случае ранения, пореза или даже царапины эти небольшие элементы склеиваются друг с другом и формируют своеобразную пробку (кровяной сгусток), которая останавливает кровотечение и препятствует попаданию болезнетворных микробов в сосудистое русло. Кроме того, тромбоциты стимулируют заживление поврежденных тканей, питают внутреннюю выстилку кровеносных сосудов, нейтрализуют бактерии и другие вредные агенты. Он бывает физиологическим и может ненадолго возникнуть практически у любого человека, например, во время беременности, после тяжелой физической нагрузки, психоэмоционального перенапряжения и т.д. Однако во всех этих случаях повышение тромбоцитов незначительное и не должно вызывать опасения. И совсем другое дело, когда их содержание превышает норму в полтора–два и более раза — такой тромбоцитоз считается патологическим и наблюдается при опухолях кроветворной ткани, амилоидозе, инфекционных заболеваниях и неинфекционных воспалительных процессах (обострении ревматизма, остеомиелите, туберкулезе), после удаления селезенки, при чрезмерном обезвоживании и т.д. Специфические симптомы тромбоцитоза отсутствуют, поскольку он сам является следствием того или иного заболевания. Соответственно следует не бороться с повышением уровня тромбоцитов, а лечить конкретный недуг.
Он бывает физиологическим и может ненадолго возникнуть практически у любого человека, например, во время беременности, после тяжелой физической нагрузки, психоэмоционального перенапряжения и т.д. Однако во всех этих случаях повышение тромбоцитов незначительное и не должно вызывать опасения. И совсем другое дело, когда их содержание превышает норму в полтора–два и более раза — такой тромбоцитоз считается патологическим и наблюдается при опухолях кроветворной ткани, амилоидозе, инфекционных заболеваниях и неинфекционных воспалительных процессах (обострении ревматизма, остеомиелите, туберкулезе), после удаления селезенки, при чрезмерном обезвоживании и т.д. Специфические симптомы тромбоцитоза отсутствуют, поскольку он сам является следствием того или иного заболевания. Соответственно следует не бороться с повышением уровня тромбоцитов, а лечить конкретный недуг. ). Все это служит сигналом для обращения за медицинской помощью. Низкая концентрация тромбоцитов в крови может быть признаком весьма серьезных заболеваний — злокачественных опухолей, аутоиммунных процессов, лучевой болезни, затянувшейся инфекции (гриппа, кори, краснухи, ветряной оспы) и т.д. Лекарства — антибиотики, мочегонные, седативные и антидиабетические — также могут привести к снижению уровня тромбоцитов.
). Все это служит сигналом для обращения за медицинской помощью. Низкая концентрация тромбоцитов в крови может быть признаком весьма серьезных заболеваний — злокачественных опухолей, аутоиммунных процессов, лучевой болезни, затянувшейся инфекции (гриппа, кори, краснухи, ветряной оспы) и т.д. Лекарства — антибиотики, мочегонные, седативные и антидиабетические — также могут привести к снижению уровня тромбоцитов.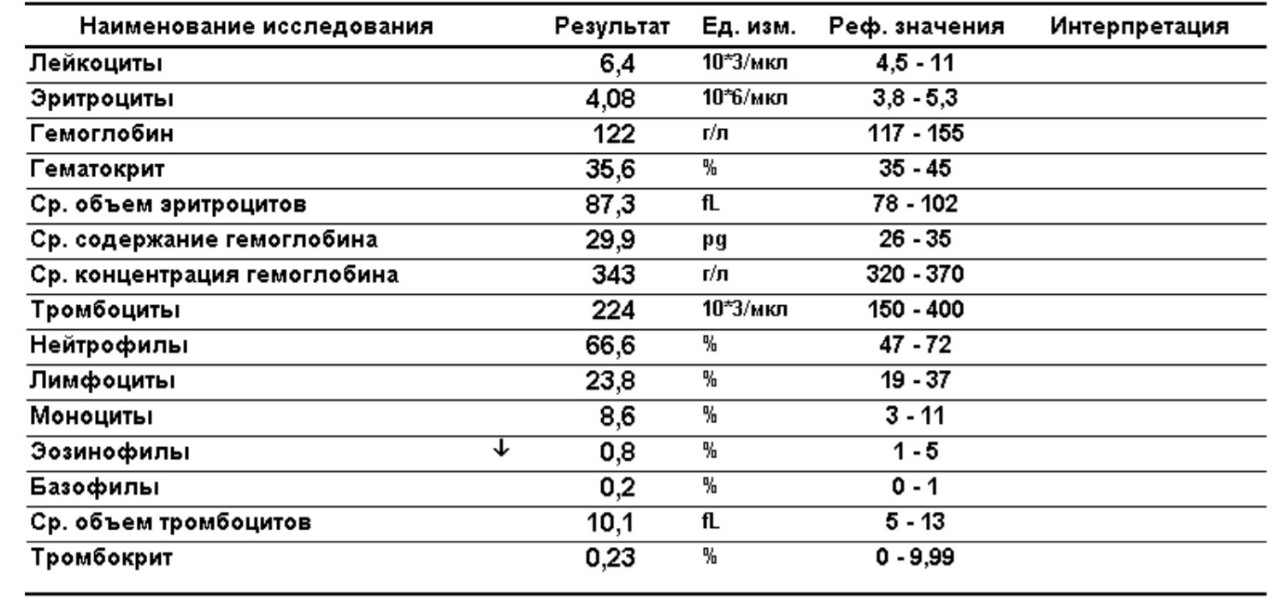 Воспаляются легкие, а в некоторых случаях — сердечная мышца или почки. Очень часто повреждаются мелкие кровеносные сосуды и повышается свертываемость крови, что, в свою очередь, может привести к образованию тромбов в легких. Именно это в большинстве случаев и становится непосредственной причиной смерти при COVID-19.
Воспаляются легкие, а в некоторых случаях — сердечная мышца или почки. Очень часто повреждаются мелкие кровеносные сосуды и повышается свертываемость крови, что, в свою очередь, может привести к образованию тромбов в легких. Именно это в большинстве случаев и становится непосредственной причиной смерти при COVID-19.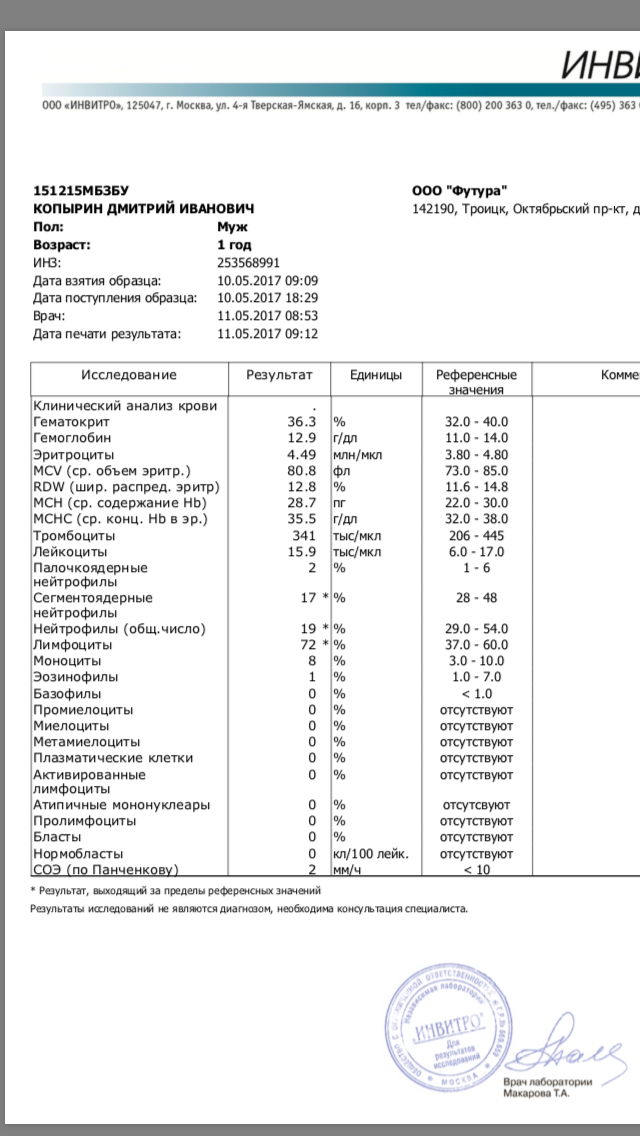 )», — рассказывает один из соавторов исследования, сотрудник Института клинической молекулярной биологии Кильского университета Флориан Тран (Florian Tran). Это может указывать на то, что мегакариоциты вызывают чрезмерное свертывание крови, что крайне опасно, поясняет он.
)», — рассказывает один из соавторов исследования, сотрудник Института клинической молекулярной биологии Кильского университета Флориан Тран (Florian Tran). Это может указывать на то, что мегакариоциты вызывают чрезмерное свертывание крови, что крайне опасно, поясняет он.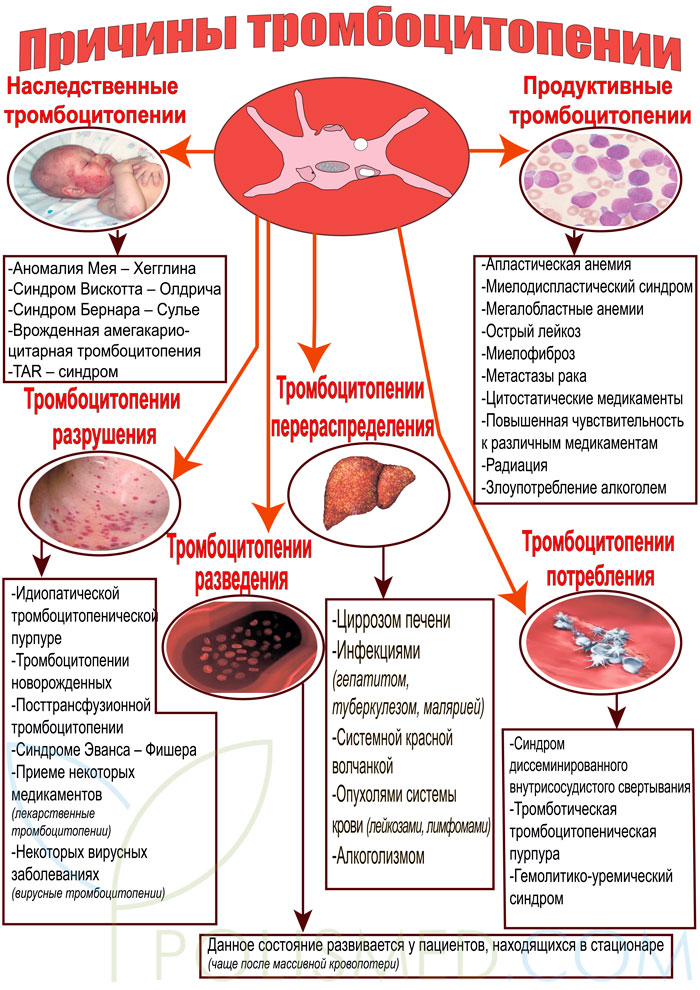
 В Рождество в большинстве земель, пока — кроме Берлина, запланированы послабления: до 10 человек. Отели для туристов закрыты. На фотографии — пустой причал на курорте Зеллин на острове Рюген.
В Рождество в большинстве земель, пока — кроме Берлина, запланированы послабления: до 10 человек. Отели для туристов закрыты. На фотографии — пустой причал на курорте Зеллин на острове Рюген.
 Обсуждая дополнительные меры при продлении локдауна, власти решили в декабре ограничить количество посетителей магазинов — в зависимости от торговой площади. Эту меру многие встретили скептически, опасаясь очередей на входах.
Обсуждая дополнительные меры при продлении локдауна, власти решили в декабре ограничить количество посетителей магазинов — в зависимости от торговой площади. Эту меру многие встретили скептически, опасаясь очередей на входах. Фотографии
Фотографии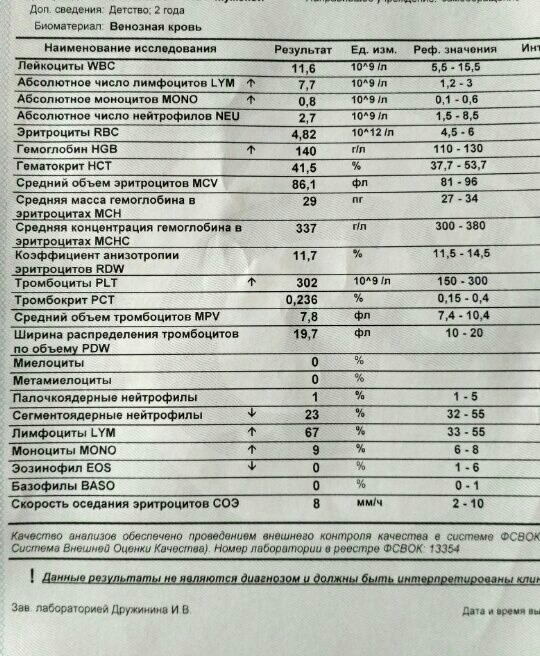
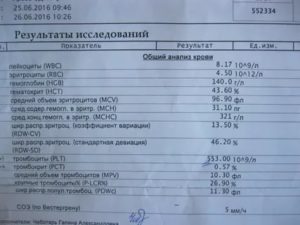
 В этой школе в Майнце испытывают новую систему вентиляции.
В этой школе в Майнце испытывают новую систему вентиляции.